深圳地区桂花夏冬季光合生理特性的比较
2012-01-04王定跃王贤荣刘永金
王定跃,曹 婷,王贤荣*,刘永金,徐 滔
(1.深圳市公园管理中心,广东 深圳 518000;2.南京林业大学森林资源与环境学院,江苏 南京 210037;3.深圳市梧桐山风景区管理处,广东 深圳 518000)
桂花(Osmanthusfragrans)为木犀科木犀属植物,是我国的传统名花,在我国的栽培历史已有2 500余年[1].桂花是寿命长、病虫害少、适应性广的常绿阔叶观赏树木,在园林和庭院的绿化、美化、香化及装饰盆景上有特殊地位[2].现在在秦岭淮河流域以南至南岭以北的广大地区均有大量的栽培,并形成了江苏苏州、四川成都、湖北咸宁、浙江杭州和广西桂林历史上著名的5大产区,而且一些新的桂花产区也正日渐发展起来[3].桂花的自然分布多为立地条件较差且其他常绿阔叶乔木难以长成大树的地段[4-6].随着现代社会城市化的发展,人们对于绿化要求也越来越高,而桂花四季常绿,在色,香,形上更是常绿树种一个代表,所以在各地都争相利用桂花这一绝佳的绿化资源.本文通过测定4个桂花品种在夏季和冬季的一系列光合作用指标,研究其净光合速率与蒸腾速率日变化特征及生理生态因子的影响,以期科学掌握光合生理,为桂花以及其他园林绿化植物的引种栽培和管理提供一个基础的参考.
1 材料和方法
1.1 材料
试验在深圳市梧桐山仙桐体育公园草坪上进行,供试材料均为2010年11月初从浙江省金华市引进的株高在1~2 m的桂花样本,分别为日香桂(O.fragrans‘Rixianggui’)、小叶佛顶珠(O.fragrans‘Xiaoye Fodingzhu’)、晚银桂(O.fragrans‘Wan Yingui’)、万点金(O.fragrans‘Wandian Jin’).样本采取统一化的水肥管理.
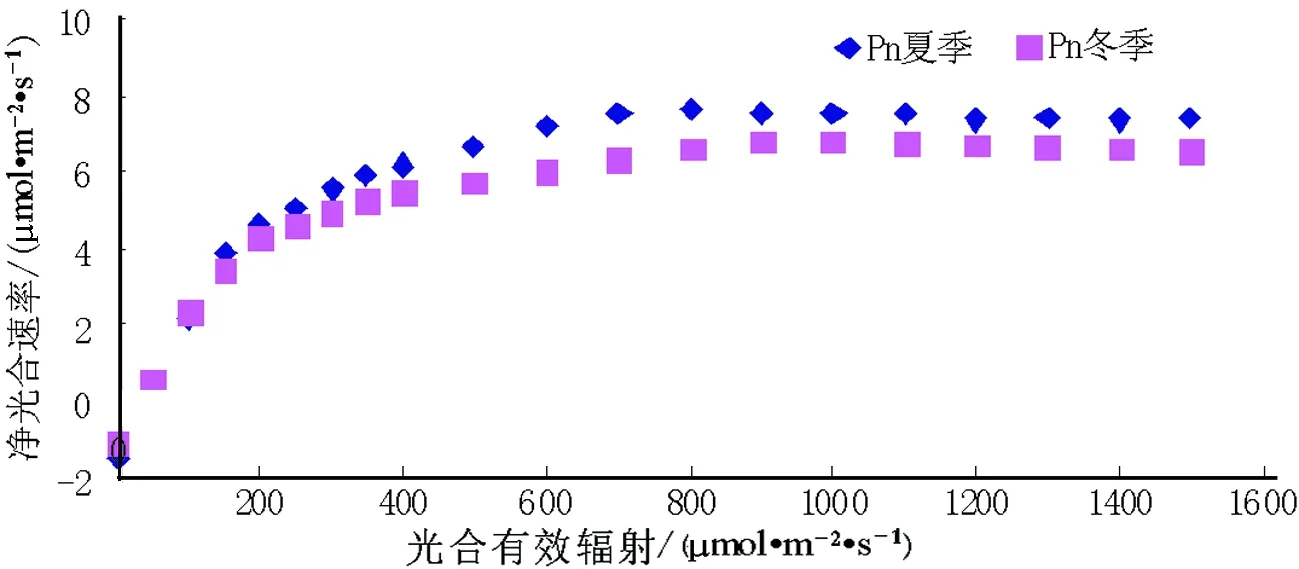
图1 夏季和冬季桂花光响应散点图
1.2 方法
试验使用LI-6400光合仪,在2011年8月中旬、12月中旬晴朗的天气进行光合测定,选取3株特定桂花枝条顶端的第1片完全伸展的当年生叶片,共3片.在8:00~18:00之间,每隔1 h对4个桂花品种叶片进行日变化各项参数测定,包括蒸腾速率(Tr)、净光合速率(Pn)、光合有效辐射 PAR、胞间CO2浓度(Ci)等.数据采用对应时刻数据的平均值,图表使用Excel 2003、SPSS17.0软件进行绘制并加以统计分析.
2 结果与分析
2.1 夏季和冬季桂花的光响应曲线分析比较
植物叶片的光饱和点与光补偿点反映了植物对光照条件的要求,分别体现了对强光和弱光的利用能力,依据植物对光环境的适应性,将植物划分为专性阳生植物、耐阴植物(兼性阳生、阴生植物)和专性阴生植物[8].实验利用LED人工光源调节光合有效辐射(PAR)强度,从0 μmol·m-2·s-1逐渐增加至2 000 μmol·m-2·s-1,测定小叶佛顶珠叶片净光合速率(Pn),并将Pn和PAR的关系绘制成曲线图(图1).

表1 夏季和冬季桂花叶片光合作用参数
由图1可以看出在夏季和冬季,当光照强度较弱时(0~200 μmol·m-2·s-1),小叶佛顶珠的Pn随着PAR的增强而呈正比例增大,Pn与PAR呈现一个线性比例阶段,本文对低PAR下的Pn进行线性回归,计算出小叶佛顶珠的光补偿点和表观量子效率(表1).当光照强度>400 μmol·m-2·s-1时,Pn随着PAR的增加而缓缓增大,但是当光照强度>1000 μmol·m-2·s-1时,Pn随着PAR的增强而呈现缓慢下降趋势.在夏季,小叶佛顶珠的光饱和点为966 μmol·m-2·s-1,光补偿点为 38 μmol·m-2·s-1,最大净光合速率为7.98 μmol·m-2·s-1;在冬季,小叶佛顶珠的光饱和点为1 028 μmol·m-2·s-1,光补偿点为30 μmol·m-2·s-1,最大净光合速率为 6.89 μmol·m-2·s-1.通过对比可以看出,夏季光饱和点〈冬季光饱和点,相反,夏季光补偿点〉冬季光补偿点,说明小叶佛顶珠在冬季对光环境的适应区间〉夏季对光环境的适应区间.小叶佛顶珠的表观光量子效率在夏季时为0.031,冬季时为0.027,表明在夏季小叶佛顶珠对光能利用效率要高于冬季.
2.2 夏季和冬季桂花光合日变化的动态比较
2.2.1 夏季和冬季环境因子日变化动态比较 从图2-3可知桂花在夏季和冬季所处的环境因子日变化均呈现一定规律的动态变化.光合有效辐射(PAR)日变化曲线在2个季节均呈现单峰型,夏季PAR强于冬季,夏季PAR的最大值为1 739 μmol·m-2·s-1,冬季PAR的最大值为742 μmol·m-2·s-1.大气温度(Ta)早晚低、午间高,日变化也呈单峰型,但是Ta在白天变化趋势缓慢.夏季Ta在整个白天都处于较高状态,最低温度是早上8:00左右出现的26℃,最高温度是中午13:00出现的37℃;冬季最高温度出现在午后14:00,只有19℃.相对湿度(RH)日变化表现为早晚高、午间低的变化趋势,在中午13:00左右,RH达到最低值,夏季RH高于冬季,夏季最低RH为22%左右,冬季最低RH只有15%左右.

图2 光合有效辐射日变化曲线 图3 温度和相对湿度日变化曲线
2.2.2 夏季和冬季桂花净光合速率和蒸腾速率日变化 植物在自然状态下,叶片的净光合速率(Pn)和蒸腾速率(Tr)日变化一般有三种情况:单峰型、非规则型和双峰型,当物种、生长阶段和天气状况等不同的时候,光合日变化动态呈现各种不同的变化类型[9].
由图4可以看出在夏季,桂花的Pn日变化曲线呈双峰曲线,明显的“午休”现象出现在光照最强、温度最高的正午时间,这与上面提到的夏季PAR日变化动态(图2)相一致.而不同品种之间,Pn存在一定的差异,其中晚银桂、万点金在一天中Pn要比其他品种大.日香桂、小叶佛顶珠、晚银桂、万点金4个桂花品种Pn第一峰值均出现在9:00,第一峰值分别为5.47、4.37、7.94、7.22 μmol·m-2·s-1,第二峰值均出现在15:00,第二峰值分别为2.15、2.29、2.89、3.29 μmol·m-2·s-1,两个峰值差异很大.由图5可以看出晚银桂、万点金的Tr曲线均是不规则的,但是最大值都出现在9:00,其值分别为3.88、3.44 mmol·m-2·s-1;日香桂、小叶佛顶珠的Tr曲线呈不明显的双峰曲线,第一峰值均出现在9:00,峰值分别为3.37、2.91 mmol·m-2·s-1、第二峰值均出现在15:00,峰值分别为2.64、2.77 mmol·m-2·s-1,两个峰值相差不大.
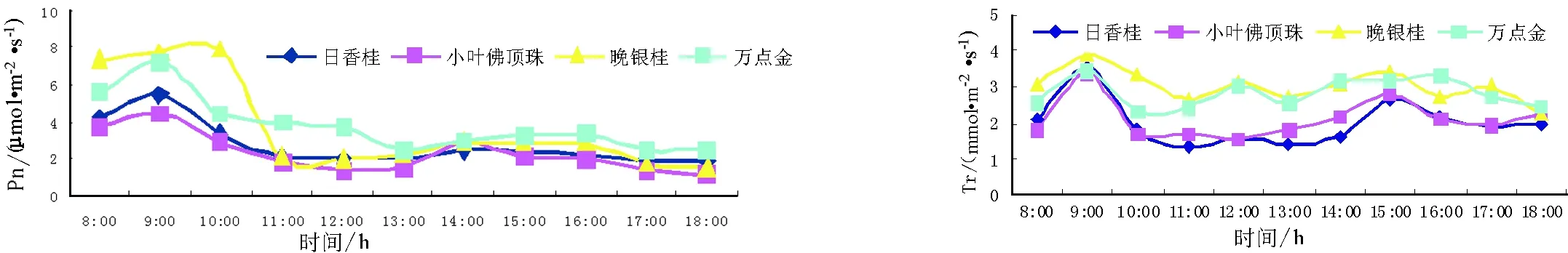
图4 8月中旬不同桂花品种净光合速率日变化 图5 8月中旬不同桂花品种蒸腾速率日变化
由图6可以看出在冬季,4个桂花品种的Pn的日变化曲线呈单峰曲线,在早晨8:00时,Pn为负值,随后缓慢上升,从9:00开始迅速增大,在正午12:00-13:00达到最大值,分别为2.66、2.48、3.77、3.87 μmol·m-2·s-1,随后迅速下降,从下午17:00开始,已经有部分品种Pn为负值,这可能是因为冬季下午光照强度和大气温度均太低,不能够满足桂花进行正常的营养生长.由图7可以看出4个桂花品种Tr总体表现为中午高,早晨和傍晚低,与Ta的日变化动态(图3)有一定的相似性,Tr总体为单峰曲线,从9:00开始迅速增大,在14:00达到最大值,分别为0.9、0.8、1.04、0.99 mmol·m-2·s-1,在冬季,桂花Tr总体变化趋势与图6中Pn结果是相一致的.

图6 12月中旬不同桂花品种净光合速率日变化 图7 12月中旬不同桂花品种蒸腾速率日变化
2.3 夏季和冬季桂花净光合速率与生理生态因子的回归分析
利用8月份测定的桂花净光合速率(Pn)和蒸腾速率(Tr)及其影响因子的数据,以Pn为应变量,以气孔导度(Cond)、胞间 CO2浓度(Ci)、饱和蒸汽压亏缺(VPD)等生理生态因子为自变量,采用SPSS17.0进行统计分析,以日香桂为例说明逐步多元回归分析过程.
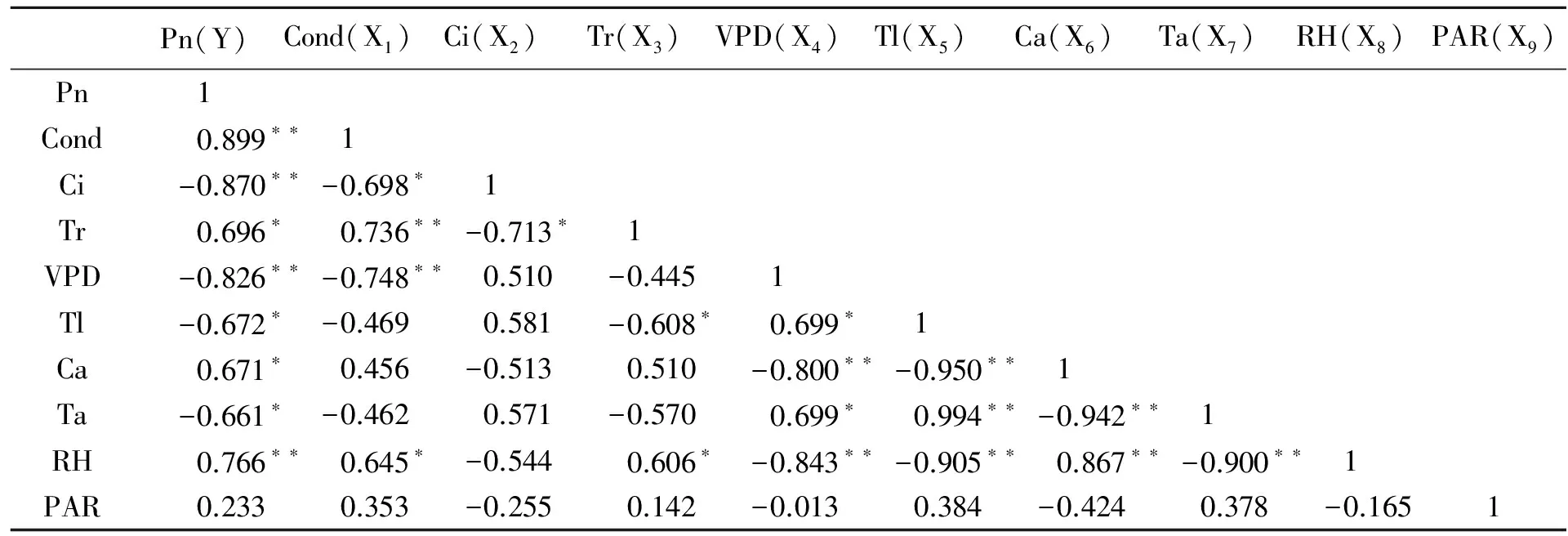
表2 日香桂净光合速率与生理生态因子的相关性分析
注:**表示在 0.01 水平上有显著意义,*表示在 0.05 水平上有显著意义.
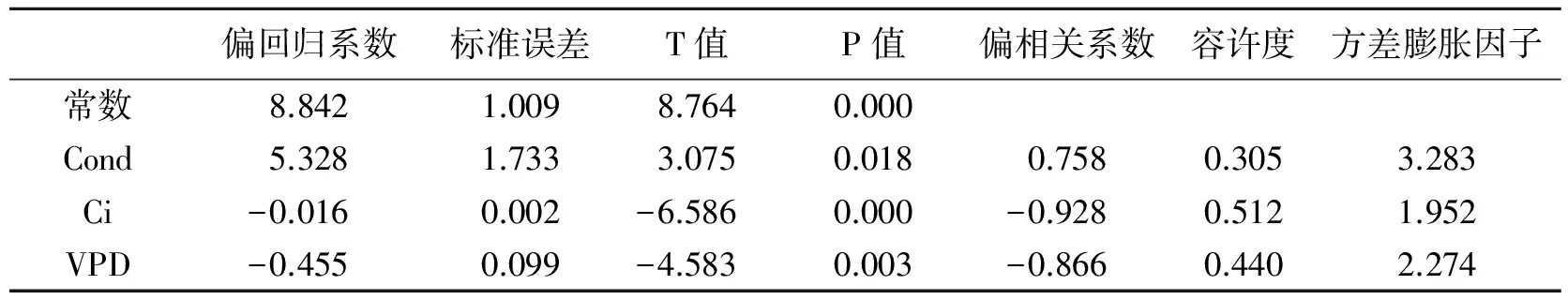
表3 回归系数分析表
由表2可以看出,日香桂叶片Pn与Cond、RH呈极显著正相关,与Tr、大气CO2浓度(Ca)呈显著正相关;与Ci、VPD呈极显著负相关,与叶温(Tl)、Ta呈显著负相关;与PAR无显著相关性.同时可以看出各个因子之间也存在复杂的相关性,为了进一步确定这些影响因子对Pn的影响程度,将日香桂的Pn与影响因子进行逐步回归,回归系数见表3,从表3中得出Pn与生理生态因子的回归方程:Y=8.842+5.328X1-0.016X2-0.455X4,R=0.990;方程明显可以看出,日香桂叶片Pn主要受Cond、Ci、VPD的影响,而偏相关系数绝对值大小,代表着相关性程度,偏相关系数绝对值越大,Pn与该因子的相关性程度就越高,因此可以看出Ci对日香桂Pn影响最大,其次是VPD和Cond.引种到深圳的不同桂花品种在夏季和冬季Pn与影响因子的逐步回归分析与日香桂相同,现将回归方程及影响因子排序结果列于表4.
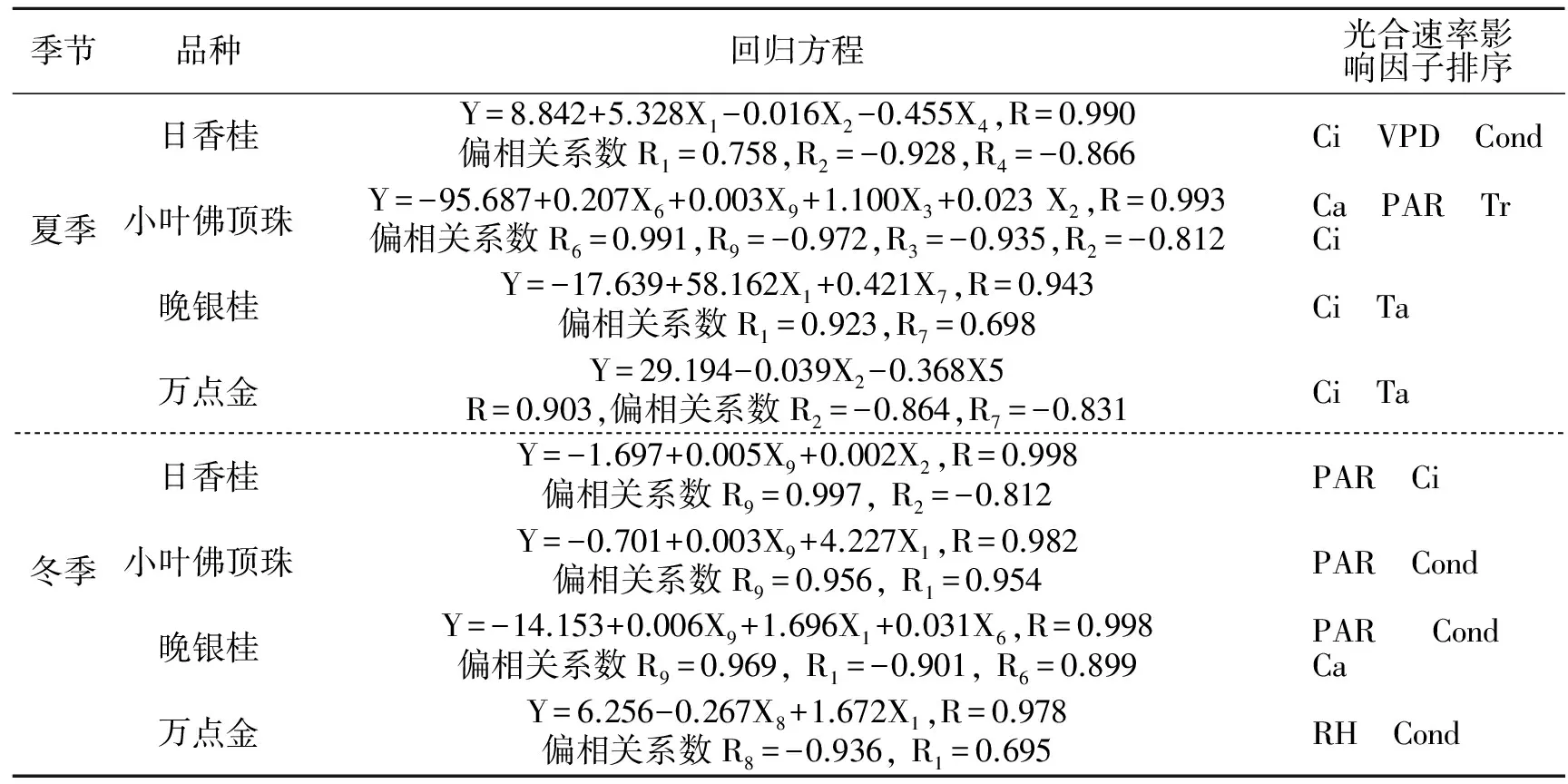
表4 逐步回归方程及基于偏相关系数的影响因子排序
从表4中可以看出桂花的Pn主要受生理生态因子的协调作用,不同桂花品种和不同季节Pn受生理生态因子的影响程度是不同的.Ci、Cond、PAR、Ta和VPD是影响4个桂花品种Pn程度较大的几个因子,其中Ci和Ta在夏季的影响程度尤为突出,PAR在冬季的影响最为明显;而在同一个季节里,影响4个桂花品种Pn的生理生态因子存在着相似性和差异性,这正说明了不同桂花品种之间,在遗传上存在一定的共性和差异,这与黄新[10]对于不同牡丹品种的研究结果一致.
3 结论与讨论
分析研究植物的光合作用,对植物生产力和产量具有一定的理论实践意义.有研究表明,植物在遭遇高温、干旱等不利胁迫时,植物本身会做出一系列反应措施,如气孔关闭、光合酶活性降低、光呼吸升高等,从而引发光合“午休”现象[11].那么根据Farquhar和Sharkey提出的气孔限制值(Ls)的分析方法,当Pn下降的同时,Ci降低,但Ls升高,表明Pn降低主要是因为气孔关闭引起的;而如果Pn下降的同时,Ci上升,但Ls下降,表明Pn降低主要是由于非气孔因素引起的[12].本研究中,在夏季,桂花出现明显的光合“午休”现象,是因为8月份正午温度过高,Tr加剧,造成气孔的关闭,Ci降低,从而降低了植物的光合作用强度,说明Cond降低引起的光合原料短缺和光合产物的积累是引起 Pn降低的主要原因,这与靳甜甜[13]、闫年[14]等人研究结果一致.在冬季,桂花不存在光合“午休”现象,是因为桂花要充分利用正午的光温条件,增大对光照的适应区间,来抵抗低温带来的不良影响.
影响桂花光合生理产生变化的因素较多,Ci、Cond、PAR、Ta和VPD是影响4个桂花品种Pn程度较大的几个因子,可以说PAR是影响桂花Pn的主导因子,其他环境因子对Pn的影响均与PAR存在一定的关系,Ta和RH受到PAR的影响而有所升降,再对Pn产生影响,所以环境因子和生理因子对桂花光合作用的影响错综复杂,它们之间还会产生复杂的连锁反应,共同影响着桂花的营养生长,这与Constable[15]等人研究结果一致.因此必须进行全面地分析,才能综合把握它们的影响,为将来桂花品种栽培管理及引种环境选择提供一定的理论依据.
[1] 向其柏,刘玉莲.中国桂花品种图志[M].杭州:浙江科学技术出版社,2008:18.
[2] 聂谷华,向其柏.桂花研究现状及其存在的问题[J].九江学院学报, 2007(3):85-87.
[3] 杨庆华.桂花的地理分布及其抗冻性研究[D].南京:南京林业大学,2006.
[4] 董建文,范小明,吴东来,等.福建长汀石蜂寨景区桂花次生林群落物种数量特征[J].植物资源与环境学报,2002,11(4): 40-44.
[5] 汪德娥.庐山山南桂花结实习性及野生群落研究[J].林业科技通讯,1997(10): 18-21.
[6] 段一凡,王贤荣,梁丽丽,等.湖北咸宁桂花品种数量分类研究[J].湖北民族学院学报:自然科学版,2010,28(3):289-293.
[7] 王贤荣,袁发银,陈昕,等.湖南浏阳桂花峡桂花次生林的物种数量特征[J].南京林业大学学报, 2004(增刊): 41-45.
[8] Lambers H,Chapin III F S,Pons T L.Plant Physiological Ecology[M].2nd edn.New York:Springer-Verlag,2008:320-374.
[9] 张志平.天目杜鹃引种栽培及生理生态特性研究[D].安微:安微农业大学,2007.
[10] 黄新.牡丹引种至巢湖的主要性状及光合生理的研究[D].安徽:安徽农业大学,2006.
[11] Dias M C,Bruggemann W.Differential inhibition of photosynthesis under drought stress in Flaveria species with different degrees of development of the C4 syndrome[J].Photosynthetica,2007,45(1):75-84.
[12] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[C]//Annual Review of Plant Physiology and Plant Molecular Biology,1982,33:317-345.
[13] 靳甜甜,傅伯杰,刘国华,等.不同坡位沙棘光合日变化及其主要环境因子[J].生态学报,2011,31(7):1783-1793.
[14] 闫年,吴文友,刘桂华,等.四个杨树品系夏季光合作用日变化[J].应用研究,2011,25(2): 48-52.
[15] Constable J V H, Peffer B J,De Nicola D M.Temporal and light-based changes in carbon uptake and storage in the springephemeral Podophyllum peltatum (Berberidaceae)[J].Environmental and Experimental Botany,2007,60(1):112-120.
