崇明水仙根尖体细胞染色体的观察和核型分析
2011-12-31周永刚张冬梅罗玉兰卞黎霞
周永刚,张冬梅,鲁 琳,罗玉兰,卞黎霞
崇明水仙为石蒜科 (Am aryllidaceae)水仙属(NarcissusL.)多年生草本植物,属于中国水仙的 1个栽培类型,与漳州水仙和普陀水仙统称为中国水仙(N.tazettaL.var.chinensisRoem.),在上海市崇明县已有 400年以上的栽培历史。崇明水仙具有花香幽雅、水养期和花期长等特点,其重瓣花型以浓郁芳香著称于国内外花卉市场,可与英国玫瑰齐名。目前,崇明水仙的栽培品种很少,且品种混杂,加之品种资质退化、品质下降及病毒病发生等因素的影响,已处于几近灭绝边缘。2007年,上海市将崇明水仙列入专项拯救计划。然而,目前还没有关于崇明水仙遗传背景的系统研究和报道。
对水仙细胞学特征进行分析是明确其种及品种间差别的基础。栗田正秀[1]、Kam ae[2]和陈晓静[3]先后进行了关于中国水仙核型的研究,均认为中国水仙是同源三倍体 (2n=3x=30);然而,王瑞等[4]以漳州单瓣花水仙为材料得出的研究结果则有所不同,确定漳州单瓣花水仙的体细胞染色体呈三倍体趋势,但其核型却显示出异源三倍体的倾向。在水仙的细胞学研究方面,李懋学等[5]于 1980年对漳州、舟山和崇明3个产地的水仙进行了染色体组型和 Giem sa C-带带型的比较研究,但并没有发现品种间的明确差异。目前,有关崇明水仙不同类型的核型还未见系统的研究报道。
作者对崇明水仙细胞学研究的前处理溶液 (对二氯苯饱和溶液和 0.002mo l·L-18-羟基喹啉水溶液)及前处理时间 (8、10、12和 14 h)进行了筛选,并在细胞学水平上对重瓣花型和单瓣花型崇明水仙根尖体细胞的染色体数、核型及倍性等进行了比较分析,以期为崇明水仙优质种源的确定、茎和花型的改良、花期的调控、切实可行的栽培保种措施的制定及其推广和应用奠定研究基础。
1 材料和方法
1.1 材料
供试的重瓣花型和单瓣花型崇明水仙鳞球均来源于上海市崇明县百叶水仙花专业合作社二基地。
1.2 方法
1.2.1 前处理液及前处理时间的筛选 选取健康、直径 8 cm的 3年生水仙商品球鳞球,每种花型 10个,经低温处理解除休眠后,剥去外层褐色的干皮膜,洗净后置于培养皿中水培,每天换水 1~2次;经 3~5 d培养,待根尖长至 0.5~1.0 cm时,在上午 9:00至 10:00时采集根尖备用。
分别用对二氯苯饱和溶液和 0.002 mo l·L-18-羟基喹啉水溶液对根尖进行前处理,浸泡时间设置 4个梯度(8、10、12和 14 h),每个时间梯度重复3次。将经过前处理的根尖用蒸馏水清洗后置于卡诺固定液〔V(无水乙醇)∶V(冰乙酸)=3∶1〕中固定 24 h;再转入 1mo l·L-1HC l溶液中于 60℃恒温条件下解离 7m in,水洗后用改良石炭酸品红染色并压片;采用冰冻法揭片,待干燥后用中性树胶封片[6]。使用LeicaDM 2000-4N30系统生物显微镜(德国莱卡公司生产)观察各处理的细胞分裂情况并计算分裂指数,从而筛选出适宜的前处理液和前处理时间。
1.2.2 根尖染色体压片及观察 根据上述预实验结果,对根尖进行前处理并按照上述实验操作流程进行制片和观察。使用Leica DM 2000-4N 30系统生物显微镜进行观察,每种花型各选择 50个染色体清晰、分散良好的中期分裂相进行观察、计数和拍照。采用Adobe Photoshop CS 8.0.1图像处理软件编辑后,各选取 10张照片按李懋学等[7]的方法进行染色体核型分析。
1.3 数据处理及计算
分裂指数 =(中期细胞数/观察细胞总数)× 100%。染色体相对长度系数(index of relative length,I.R.L)组成按照 Kuo等的方法[8]进行分析,染色体相对长度计算公式为:I.R.L=(染色体长度/全组染色体总长度)×100%;臂比计算公式为:臂比=长臂长度 /短臂长度;按照 Stebbins的标准[9]进行核型分类;核型不对称系数 (asymm etrical karyotype coefficient,A s.k)按照 A rano的方法[10]计算,计算公式为:A s.k=(长臂总长/全组染色体总长)×100%。
2 结果和分析
2.1 适宜的前处理液和前处理时间
为提高崇明水仙根尖体细胞中期细胞的分裂指数,采用对二氯苯饱和溶液和 0.002mo l·L-18-羟基喹啉溶液这 2种常用于植物染色体的前处理液对根尖进行预处理,并分别设置了 8、10、12和 14 h的前处理时间。结果表明 (表 1):采用不同的前处理液进行前处理,崇明水仙根尖所需的前处理时间不同;对二氯苯饱和溶液的前处理效果优于 0.002 mol· L-18-羟基喹啉溶液,染色体处于分裂中期的细胞数较多,分裂指数最高可达 7.21%,且染色体容易分散、结构比较清晰。因此,适宜于崇明水仙细胞学实验的前处理液为对二氯苯饱和溶液,最佳前处理时间为12 h。
2.2 染色体数及倍性分析
对重瓣花型和单瓣花型崇明水仙各 50个根尖细胞的中期分裂相进行染色体计数,结果表明:重瓣花型有 46个细胞的染色体数为 30,占计数细胞总数的92%;单瓣花型有 48个细胞的染色体数为 30,占计数细胞总数的 96%。由图 1可见:根据染色体的形态、长度和长短臂比,重瓣花型和单瓣花型崇明水仙体细胞的 30条染色体都可以按照每组 3条匹配为 10组,故推测它们都是 x=10的三倍体。因此,确定崇明水仙体细胞的染色体数为 2n=3x=30。
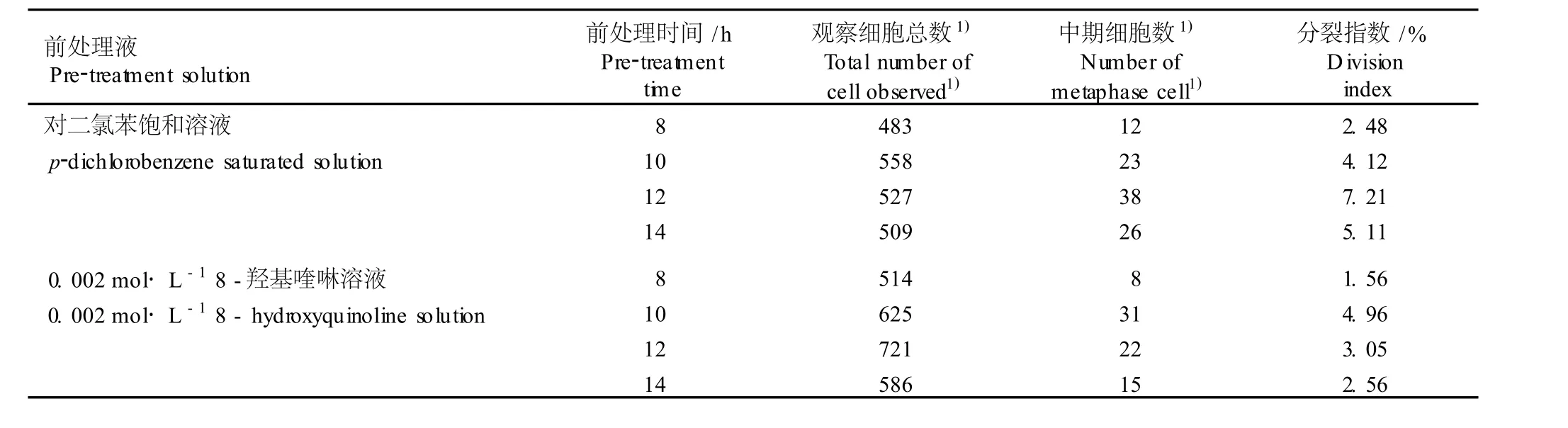
表 1 前处理液和前处理时间对崇明水仙根尖体细胞中期细胞分裂指数的影响Tab le 1 Effec t of p re-trea tm en t so lu tion and tim e on d iv ision index ofm etaphase cell in som a tic cells of root-tips of Chongm ing narc issus (N a rcissus tazetta L.var.ch inensis Roem.)
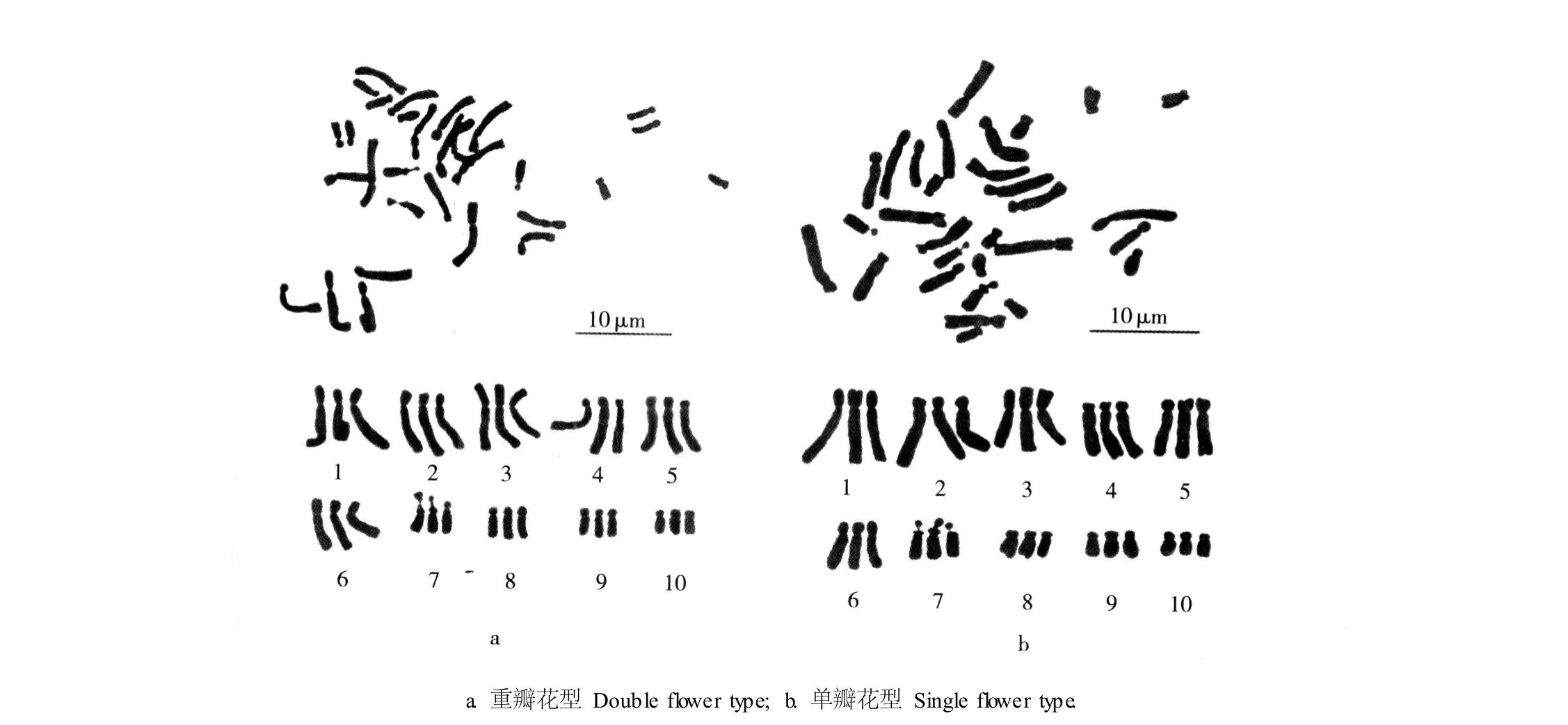
图 1 重瓣花型和单瓣花型崇明水仙根尖体细胞的染色体形态Fig.1 Chrom osom em orpho logy in som a tic cellsof root-tips of doub le and single flower types of Chongm ing narcissus(N a rcissus tazetta L.var.ch inensis Roem.)
2.3 染色体核型分析
2.3.1 重瓣花型崇明水仙染色体核型分析 根据根尖体细胞染色体的配对结果对重瓣花型崇明水仙的染色体核型参数进行统计分析,结果见表 2;根据表 2数据绘制的染色体核型模式图见图 2。
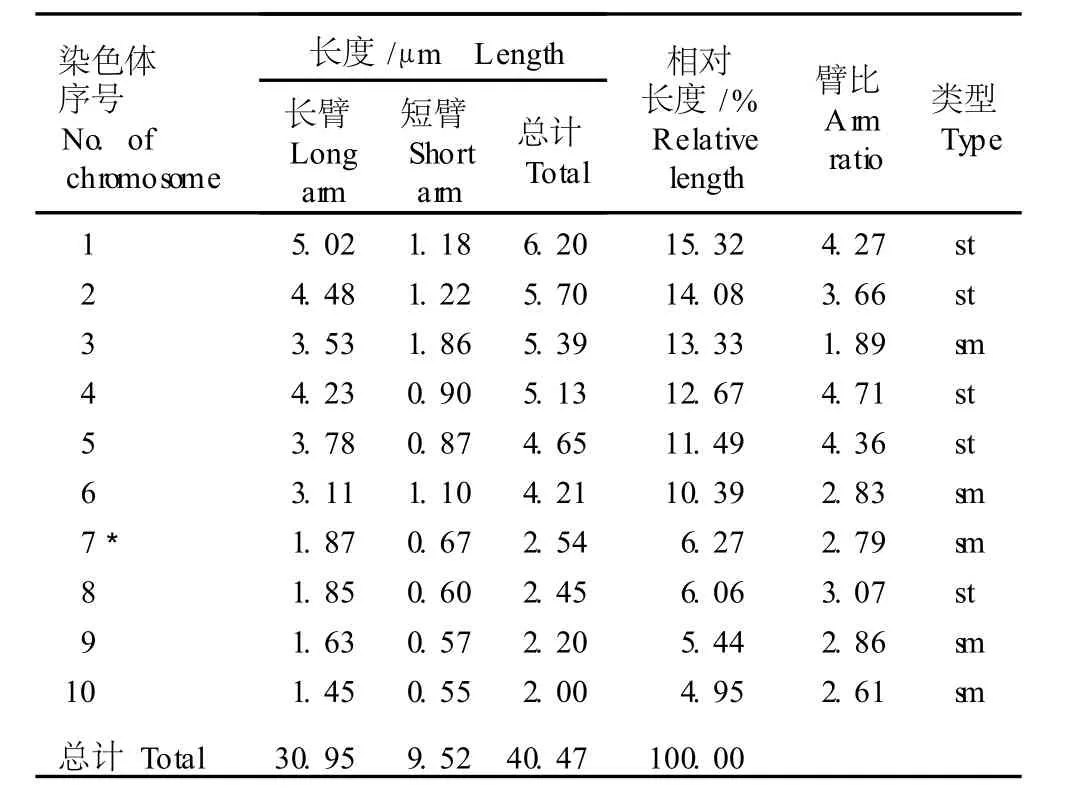
表 2 重瓣花型崇明水仙根尖体细胞染色体的核型参数1)Tab le 2 Karyotype param eters of chrom osom es in som a tic cells of root-tips of doub le flower type of Chongm ing narc issus(N a rcissus tazetta L.var.ch inensis Roem.)1)
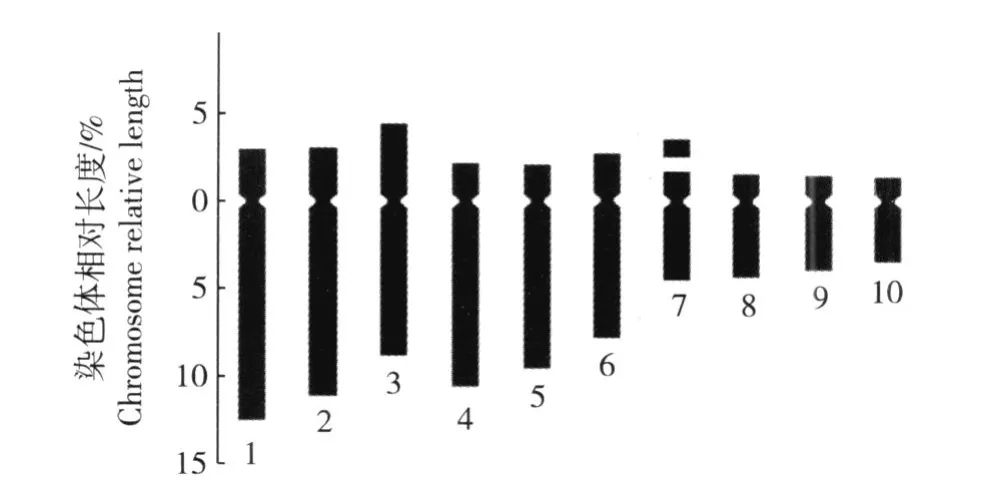
图 2 重瓣花型崇明水仙根尖体细胞染色体核型模式图F ig.2 Karyotype d iagram of chrom osom es in som a tic cells of roottips of doub le flower type of Chongm ing narcissus(N a rcissus tazetta L.var.ch inensis Roem.)
在 10组染色体中,第 3、6、7、9和 10号染色体为近中部着丝点染色体(臂比小于 3),其余染色体均为近端部着丝点染色体(臂比大于 3);在第 7号染色体上有随体,随体长度为 0.49μm。染色体组总长度为40.47μm;长臂总长度为 30.95μm,平均长度为4.05μm。在 10组染色体中第 1号染色体的总长度最长,为 6.20μm;第 10号染色体最短,为 2.00μm;最长染色体与最短染色体长度的比值为 3.10。核型不对称系数(A s.k)为 76.48%,相对长度系数组成为2n=30=12L+6M2+12S。分析结果显示:重瓣花型崇明水仙根尖体细胞染色体的核型公式为 2n=3x= 30=15st+15sm(3SAT)。
由表 2还可看出:重瓣花型崇明水仙体细胞染色体的臂比变化范围为 1.89~4.71,臂比大于 2的染色体有 9组,占全部染色体组的 90%。根据 Stebbins的核型分类标准,重瓣花型崇明水仙的体细胞的染色体核型属于“3B”型。
2.3.2 单瓣花型崇明水仙染色体核型分析 根据根尖体细胞染色体的配对结果对单瓣花型崇明水仙的染色体核型参数进行统计分析,结果见表 3;根据表3数据绘制的染色体核型模式图见图 3。
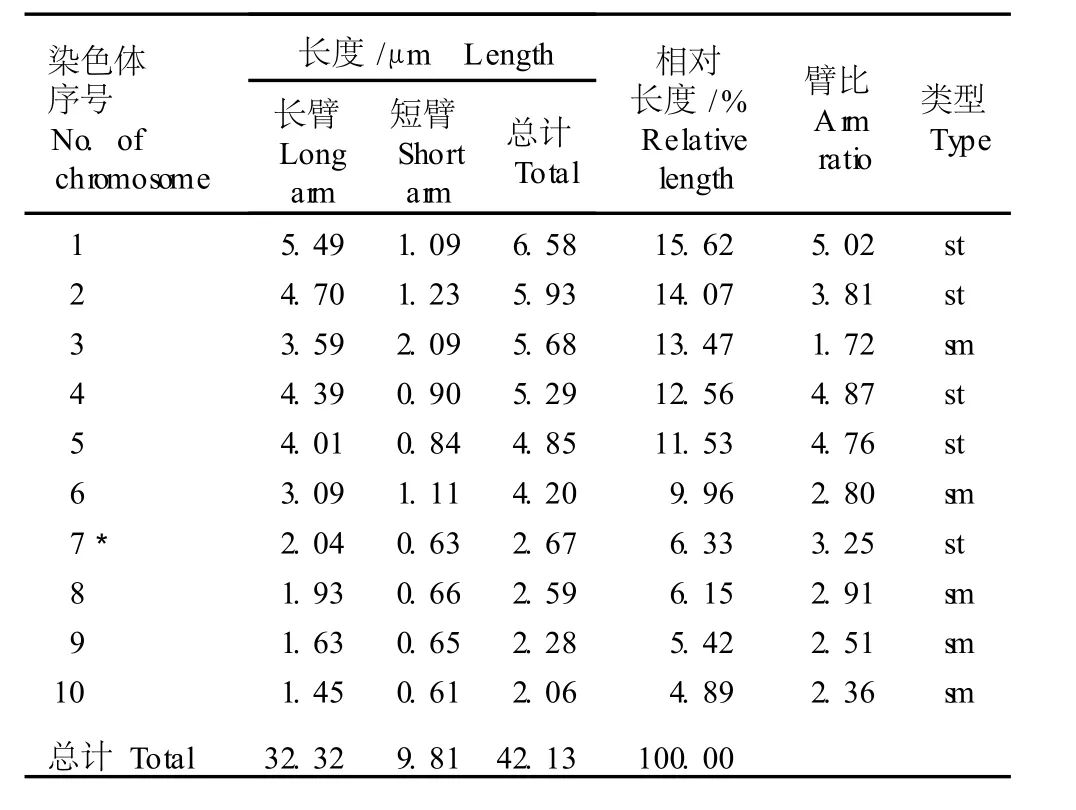
表 3 单瓣花型崇明水仙根尖体细胞染色体的核型参数1)Tab le 3 Karyotype param eters of chrom osom es in som a tic cells of root-tips of single flower type of Chongm ing narcissus(N a rcissus tazetta L.var.ch inensis Roem.)1)
在 10组染色体中,第 3、6、8、9和 10号染色体为近中部着丝点染色体(臂比小于 3),其余染色体均为近端部着丝点染色体(臂比大于 3);在第 7号染色体上有随体,随体长度为 0.52μm;染色体组总长度为42.13μm;长臂总长 32.32μm,平均长度 4.21μm。在10组染色体中第1号染色体的总长度最长,为 6.58μm;第 10号染色体最短,为 2.06μm;最长染色体与最短染色体长度的比值为 3.19。核型不对称系数(A s.k)为 76.71%,相对长度系数组成为 2n=30= 12L+3M1+3M2+12S。分析结果显示:单瓣花型崇明水仙根尖体细胞染色体的核型公式为 2n=3x=30=15st(3SAT)+15sm。
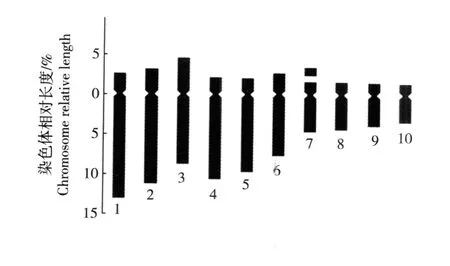
图 3 单瓣花型崇明水仙根尖体细胞染色体核型模式图F ig.3 Karyotype d iagram of chrom osom es in som a tic cells of roottips of sing le flower type of Chongm ing na rc issus(N a rcissus tazetta L. var.ch inensis Roem.)
由表 3还可以看出:单瓣花型崇明水仙根尖体细胞染色体的臂比变化范围为 1.72~5.02,臂比大于 2的染色体有 9组,占全部染色体组的 90%。根据Stebbins的核型分类标准,单瓣花型崇明水仙体细胞染色体的核型属于“3B”型。
2.3.3 重瓣花型和单瓣花型崇明水仙染色体平均核型分析 根据上述观察和测量结果可见:重瓣花型和单瓣花型崇明水仙的根尖体细胞染色体核型间的差异不明显,将二者的核型参数进行平均分析得到的结果见表 4,根据平均核型参数绘制的平均核型模式图见图4。
在 10组染色体中,第 3、6、8、9和 10号染色体为近中部着丝点染色体(臂比小于 3),其余染色体均为近端部着丝点染色体(臂比大于 3);第 7号染色体具有随体。染色体组总长度 41.28μm;长臂总长 31.61μm,平均长度 4.13μm。核型不对称系数 (A s.k)为76.57%,相对长度系数组成为 2n=30=12L+6M2+ 12S。分析结果显示:崇明水仙根尖体细胞染色体的平均核型公式为 2n=3x=30=15st(3SAT)+15sm。
由表 4还可以看出:崇明水仙根尖体细胞染色体中臂比大于 2的染色体有 9组,占全部染色体组的90%。根据 Stebbins的核型分类标准,崇明水仙体细胞染色体的核型属于“3B”型。
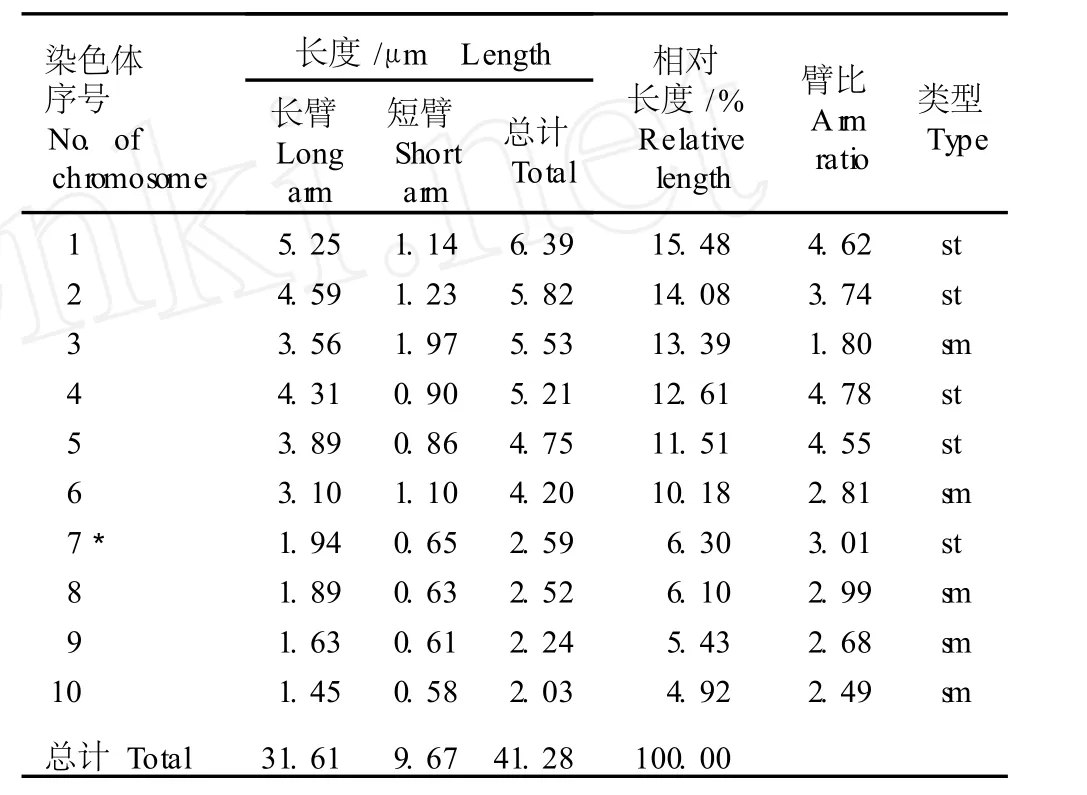
表 4 崇明水仙根尖体细胞染色体的平均核型参数1)Tab le 4 Average karyotype param eters of chrom osom es in som a tic cells of root-tips of Chongm ing narc issus(N a rcissus tazetta L.var. ch inensis Roem.)1)
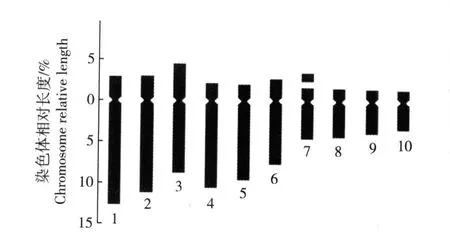
图 4 崇明水仙根尖体细胞染色体平均核型模式图F ig.4 Average karyotype d iagram of chrom osom es in som a tic cells of roo t-tips of Chongm ing narc issus(N a rcissus tazetta L.var. ch inensis Roem.)
2.3.4 重瓣花型和单瓣花型崇明水仙染色体核型的比较分析 重瓣花型和单瓣花型崇明水仙根尖体细胞染色体核型参数的比较结果见表 5。结果表明:重瓣花型和单瓣花型崇明水仙根尖体细胞染色体的核型略有差异。相同点为:都属于典型不对称的二型核型,染色体数和倍性相同,核型类型同属于“3B”型,随体均出现在第 7号染色体的短臂上,臂比大于 2的染色体组数占全部染色体组的 90%。差异在于:重瓣花型的第 7号染色体为 sm型、第 8号为 st型,而单瓣花型的第 7号染色体为 st型、第 8号为 sm型;前者的核型不对称系数(A s.k)略小于后者;前者的相对长度系数为 12L+6M2+12S,后者的相对长度系数为 12L +3M1+3M2+12S;前者的最长染色体与最短染色体长度的比值略小于后者。综合李懋学等[5]的研究结果,可以认为重瓣花型和单瓣花型崇明水仙具有同一起源,二者在形态特征方面的差异可能是长期进行无性繁殖和栽培选择所形成的基因型和生态型的差异。
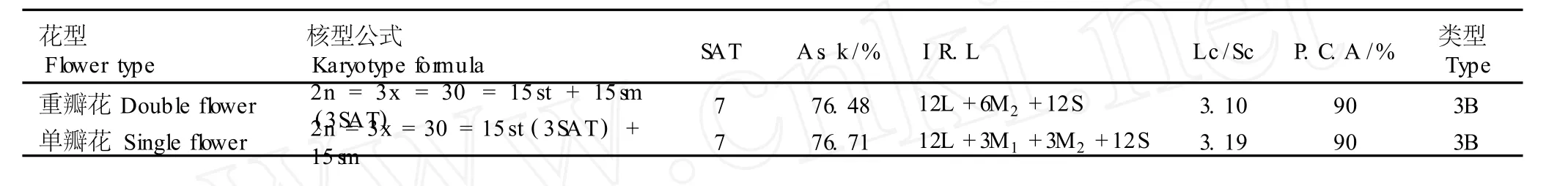
表 5 重瓣花型和单瓣花型崇明水仙根尖体细胞染色体核型参数的比较1)Tab le 5 Com par ison of karyotype param eters of chrom osom es in som a tic cells of root-tips of doub le and single flower types of Chongm ing narcissus(N a rcissus tazetta L.var.ch inensis Roem.)1)
3 讨论和结论
在植物细胞染色体核型分析过程中,前处理过程是使细胞分裂停止在中期阶段而获得较多中期分裂相的关键步骤,前处理同时还可以使染色体缩短变粗,便于观察和统计。在本实验过程中,针对不同类型水仙,前处理液和前处理时间有一定差异。前处理时间过短,染色体大部分还处于染色质状态,得不到清晰可见的染色体;前处理时间过长,造成染色体过度缩短,无法判别着丝点,也无法体现各染色体之间的差异,不能进行准确的核型分析。本实验中,在染色体制片过程中利用对二氯苯饱和溶液前处理 12 h,能够获得较多的中期分裂相细胞。但在 50个中期分裂相细胞中,能够清晰鉴别出具有 30条染色体数的细胞数未达到 100%,这可能是由于在染色体制片过程中出现重叠或断裂所致。
核型分析结果表明:重瓣花型和单瓣花型崇明水仙根尖体细胞的染色体核型基本一致,差异较小。染色体数都为 2n=3x=30,其中包括 18条长染色体和12条短染色体,短染色体中有 3条具随体,按照染色体长短和随体染色体数目进行配对可以配成 10组,平均核型公式为 2n=3x=30=15st(3SAT)+15sm。根据现有研究资料可以确定崇明水仙是三倍体植物。
水仙属植物染色体数目和倍性变化复杂:染色体基数(x)有 7、10和 11三类;除二倍体外,还存在三倍体、四倍体和六倍体;染色体数从 14~56不等[11-13]。通过细胞学观察,一些学者认为中国水仙是三倍体植物[1,5],但是也有学者持不同观点。曾沧江等[14]认为中国水仙是二倍体植物;李懋学等[5]发现崇明水仙的染色体数目 2n=30,并据此推断崇明水仙为同源三倍体植物。与王瑞[15]报道的漳州水仙染色体核型相比,崇明水仙的 10组染色体组中有多组出现 3条染色体中 2条体积形态相似并与另外一条形态差异较大的现象,尤其是第 7组具随体的 3条染色体,其中的 2条无论是长度、臂比还是随体长度都相似,而第 3条则随体明显,且染色体长度在 3条染色体中最短。因此,初步推测崇明水仙是节段异源三倍体。这一结果与朱心武等[16]、庄伟建等[17]和李广苹[18]的研究结果一致。至于这种核型差异究竟是同源三倍体染色体之间的杂合现象,还是节段异源三倍体之间的差别,需要通过分子原位杂交和分子标记等实验进行验证。
由于长期以来一直进行无性繁殖,因此水仙的后代性状相对稳定。重瓣花型和单瓣花型崇明水仙体细胞染色体的核型差异较小,但在形态特征方面则表现出一定的差异,这可能与不同生态型的差异有关。从不同生态型的崇明水仙中选择形态性状较好的类型作为优质种源进行迅速繁殖推广,可达到对崇明水仙种源进行提纯、复壮和保种的目的,具有重要理论意义。
三倍体植物属于多倍体范畴,多倍体特性不仅体现在植株速生和营养物质增多上,还体现在植株的生态适应性及抗逆性增强等方面[19]。从长期的栽培历史来看,崇明水仙具有球茎紧实、水养期及花期长、适应性强、对 SO2和 CO有较强抗性等特性,在形态和生理上初步显示出多倍体的物种优势。目前,利用秋水仙素处理植物组织器官[20]以及原生质体融合技术在多倍体育种上都取得显著效果[21]。多倍体育种可增加原细胞型不具有的新特性[22],可使花卉的花器官增大、色彩更艳丽或花期延长[23]。综上所述,基于崇明水仙所具有的三倍体遗传背景,对崇明水仙进行倍性改良具有很大的应用潜力。然而,由于崇明水仙鳞茎小、少侧脚、叶片较小、花多独枝且不易造型,因此,可以考虑采用秋水仙素处理使染色体加倍的方法诱导崇明水仙产生突变,以获得鳞茎增大、侧脚和花枝增多、花器官增大的崇明水仙品种。
不同植物都有自身最适宜的染色体倍性水平,并不是染色体数越多、倍性越高越佳。如果超过了一定的倍性限度,就会破坏植物的遗传平衡和生理平衡,从而导致许多缺陷的产生,尤其在同源多倍体中这一现象更为突出。中国水仙是异源三倍体植物,其多倍体的诱导与二倍体的纯合加倍有所区别,诱导多倍体时极易出现嵌合体,且目前已有报道的是诱导出鳞茎球大于三倍体的六倍体[15]。因此,对于崇明水仙多倍体的诱导及其最适倍性有待于进一步深入研究。
[1]栗田正秀.水仙属的细胞学研究 I.NarcissustazettaL.的变种及园艺品种的核型[J].育种杂志,1955,5(1):23-26.
[2]KamaeM.Fundam ental studies on the breeding of JapaneseNarcissus(N.tazettaL.var.chinensisRoem.)[J].Bulletin of the Faculty of Education of KobeUniversity,1976,40:61-76.
[3]陈晓静.福建 3个产地水仙的核型分析[J].植物资源与环境学报,2004,13(4):28-31.
[4]王 瑞,陈林姣,赵玉辉,等.中国水仙的核型分析和小孢子发生中的细胞学研究[J].细胞生物学杂志,2007,29(1):140-146.
[5]李懋学,陈定慧,王莲英.中国水仙的染色体组型和 Giem sa C-带之带型研究[J].园艺学报,1980,7(2):29-37.
[6]朱 澂.植物染色体及染色体技术[M].北京:科学出版社, 1982:42-98.
[7]李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
[8]Kuo SR,W ang T T,Huang T C.Karyotype analysisof some Formosan gymnosperm s[J].Taiwania,1972,17(1):66-80.
[9]Stebbins G L.Chromosomal Evolution in H igher Plants[M]. London:Edward A rnold,1971:85-104.
[10]A rano H.Cytological studies in subfam ily Carduoideae(Compositae)of Japan.Ⅸ[J].BotanicalM agazine of Tokyo,1963,76: 32-39.
[11]Darlington CD,W ylieA P.ChromosomesA tlasof Flowering Plants [M].2nd ed.London:GeorgeA llen and Unw in L td.,1955:20-26.
[12]吕柳新,陈晓静,余小玲,等.水仙品种资源的育种基础研究I.多花水仙若干品种类型的细胞学研究[J].福建农学院学报,1989,18(1):31-36.
[13]B randham P E.Evolution of polyp loidy in cultivatedNarcissussubgenusNarcissus[J].Genetiea,1986,68(3):161-167.
[14]曾沧江,陈勤娘.福建漳州水仙花的染色体数目及命名研究[J].植物研究,1984,4(4):159-163.
[15]王 瑞.中国水仙的细胞学观察和六倍体诱导[D].厦门:厦门大学生命科学学院,2007:25-42.
[16]朱心武,徐晋麟,杨光锐.中国水仙的核型研究[J].武汉植物学研究,1986,4(2):119-122.
[17]庄伟建,林治良,苏金为.中国水仙两个地方品种的核型和Giem sa C带的研究[J].福建农业学报,1999,14(1):51-57.
[18]李广苹.漳州水仙随体单染色体文库的构建及其AFLP标记的研究[D].福州:福建农林大学园林学院,2009:17-24.
[19]罗玉兰,张冬梅,蔡 润.红刺玫染色体的观察和核型分析[J].山东林业科技,2007(3):1-2.
[20]KapoorM L,Sharma V K.Experimentally synthesized allotetrap loids inEucalyptus[J].Silvae Genetica,1985,34(1):19-22.
[21]雷加军,吴禄平,代汉萍,等.草莓茎尖染色体加倍研究[J].园艺学报,1999,26(1):13-18.
[22]Lumaret R.Adap tives strategies and p loidy levels[J].Acta Oecologica-Oecologia Plantarum,1988,9:83-93.
[23]曾正明,郭启高.植物多倍体形成途径及机制[J].绵阳经济技术高等专科学校学报,2000,17(4):1-4.
