悬尾应激对小鼠空间记忆及其反转学习的损伤效应
2011-12-25王晓琴王功伍段秋燕滕仕通
王晓琴, 王功伍, 段秋燕, 滕仕通
(1. 云南师范大学 体育学院,云南 昆明 650500; 2. 云南师范大学 生命科学学院行为与认知神经生物学实验室,云南 昆明 650500; 3. 生物能源持续开发利用教育部工程研究中心和云南省生物质能与环境生物技术重点实验室,云南 昆明 650500)
悬尾应激对小鼠空间记忆及其反转学习的损伤效应
王晓琴1, 王功伍2,3,*, 段秋燕2, 滕仕通2
(1.云南师范大学 体育学院,云南 昆明650500; 2.云南师范大学 生命科学学院行为与认知神经生物学实验室,云南 昆明650500; 3.生物能源持续开发利用教育部工程研究中心和云南省生物质能与环境生物技术重点实验室,云南 昆明650500)
该文探讨了连续数天短时悬尾应激对小鼠十字迷宫空间记忆及其反转学习的作用。81只成年雄性昆明小鼠被分为4大组:绝对空间记忆获得组及巩固组; 相对空间记忆获得组及巩固组。每大组又分为悬尾组(每天训练前或训练后立即接受悬尾处理20 min)和对照组。结果表明, 在空间记忆训练初期, 各对照组和悬尾组动物正确反应率无明显差异, 均在机遇水平; 随着训练天数的增加, 对照组成绩显著提高, 当其正确反应率达到80%时, 悬尾组正确反应率仍处于或略高于机遇水平, 两组间差异显著(P< 0.01); 在反转学习中, 悬尾组正确反应率也显著低于对照组(P< 0.01)。这些表明,悬尾应激可显著损伤小鼠的空间记忆及其反转学习的获得和巩固, 其中相对空间记忆及其反转学习的巩固受损尤为严重。
悬尾应激; 空间记忆; 反转学习; 获得; 巩固; KM小鼠
悬尾(tail suspension)处理可使动物处于无法摆脱的应激状态, 易出现行为绝望, 与人类的抑郁症表现类似, 并对很多抗抑郁药敏感, 故与强迫游泳一起成为筛选抗抑郁药物最常用的动物行为模型(Cryan et al, 2005; Seong et al, 2002)。悬尾处理同其他应激方式一样, 可通过各个感觉通道和下丘脑-垂体-肾上腺皮质轴(hypothalamus- pituitary- adrenal cortex axis, HPA轴)产生广泛的心理、生理应激效应(Cui et al, 2008), 但悬尾处理有其独特之处, 即可使动物的体液(尤其是血液)头向分布。人在大部分时间里处于坐、立位状态, 循环系统已经适应了在地球重力作用下, 体液脚向集中的情形, 并升高血压以保证头部血液的供应。航天员进入太空后, 处于失重状态, 其所受重力接近于零, 此时,1.5~2L的体液会迅速从下肢转移到头胸部, 并引发一系列的感觉神经活动改变和心血管反应(Wu & Shen, 2000)。动物在悬尾后头部下垂, 躯体受重力作用方向改变, 导致体液向头部转移, 故悬尾动物体液分布改变状况与处于失重状态下的人相似, 可作为研究人体失重的动物模型 (Chen et al, 1993; Morey-Holton & Globus, 2002; Morey-Holton & Wronski, 1981)。虽然空间飞行可直接提供失重环境, 但又伴随噪声、机械振动、宇宙射线、人际交往等因素的干扰。因此,很难研究失重等单一影响因素的效应。悬尾处理为此提供了可行的地面失重模型。
单次悬尾处理20 min可模拟急性失重, 主要通过影响动物的神经和心血管系统功能发挥其应激效应。有报告指出, 单次悬尾处理20 min可明显提升小鼠血液中糖皮质激素的水平并使之持续20 min不降低, 其情绪记忆获得在悬尾后短时受损。单独使用多巴胺D1受体拮抗剂SCH23390可损伤情绪记忆获得, 但与悬尾处理同时使用却可改善悬尾对情绪记忆获得的损伤效应, 说明多巴胺D1受体参与了悬尾应激的记忆损伤效应 (Wang et al, 2005, 2007)。但该种悬尾处理方式对空间记忆各过程的影响还未被深入系统地研究。本研究拟采用相似的悬尾处理方法(每天训练前或训练后,悬尾20 min), 探讨悬尾应激对绝对和相对空间记忆及其反转学习获得和巩固过程的影响。
1 材料和方法
1.1 实验动物及分组
81只雄性昆明(KM)小鼠, 购自成都达硕生物科技有限公司, 生产许可证编号:SCXK(川)2008-24。小鼠购入时为4周龄, 体重18~25 g。小鼠分笼饲养, 每笼(组)10~11只, 自由饮水。动物总共分4大组, 分别为绝对空间记忆获得组及巩固组和相对空间记忆获得组及巩固组。根据是否接受悬尾处理每大组动物又分为对照组及悬尾组。正式实验前适应1周, 实验者每天抚摸、抓握小鼠数次。饲养间保持12 h / 12 h的明暗周期变化(明亮期:7:00 —19:00), 室温保持在(22 ± 1) ℃。所有行为训练均在每天明亮期同一时间进行。适应期动物自由取食(标准饲料购自苏州双狮实验动物饲料科技有限公司)。正式实验时, 行为训练完成后单独定量喂食, 控制小鼠的体重为自由取食状态下的90%左右。所有实验过程均遵照国务院《实验动物管理条例》(1998)和《云南省实验动物管理条例》(2007)以及美国国立健康研究院《实验动物使用和照管指南》执行。
1.2 动物空间记忆作业
1.2.1 实验装置 所有空间记忆作业均使用一木制十字形迷宫进行。该迷宫放置于距离地面50 cm的平台上, 每臂长30 cm, 宽5 cm, 高12 cm, 四臂中央区为5 cm × 5 cm的正方形。四臂末端内侧1.5 cm处均装有直径 3 cm, 高 1 cm的圆筒状塑料食槽(图1)。迷宫正上方悬挂有两盏40 W日光灯用于照明。实验期间, 迷宫位置不变, 周围分布的门窗、实验台、实验桌和药品柜用作空间记忆参照物。
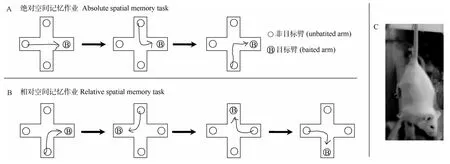
图1 十字迷宫空间记忆作业流程图Fig. 1 The flow diagram of the spatial memory tasks in plus maze
1.2.2 训练方法 根据空间信息性质的不同, 我们设计了绝对空间记忆作业和相对空间记忆作业。在绝对空间记忆作业中, 固定迷宫的一臂为目标臂(东侧臂), 其臂端食槽内放置新鲜去皮向日葵籽作为奖励, 其余三臂为起始臂, 其臂端无食(图1A)。每天训练10次(采用半随机表法确定10次训练中起始臂的顺序), 当对照组动物连续两天在作业中的正确反应率保持在80 %以上(即在10次训练中有8次以上动物能进入目标臂并获得食物奖励)时, 对照组和悬尾组动物均停止训练。然后改对侧臂为目标臂(西侧臂), 其余三臂为起始臂, 照以上方法继续进行训练, 当对照组动物在作业中达到80 %的正确反应率标准时, 两组动物均停止训练, 此即绝对空间反转学习作业。在相对空间反转学习作业中,起始臂不固定(采用半随机表法确定10次训练中起始臂的顺序), 且每次训练均须动物从起始臂出发后向右转才能到达目标臂获得食物奖励(即正确反应), 即目标臂总是起始臂的右侧臂, 起始臂改变了,目标臂也就跟着改变(图1B)。每天训练10次, 当对照组动物连续两天在作业中的正确反应率保持在80%以上时, 对照组和悬尾组动物均停止训练; 然后,改变目标臂与起始臂的相对位置(目标臂改为起始臂的左侧臂), 动物须从起始臂出发后向左转才能到达目标臂并获得食物奖励(即正确反应), 此为相对空间反转学习作业。当对照组动物在作业中达到80 %的正确反应率时, 对照组和悬尾组动物均停止训练。动物在迷宫中每完成一次作业, 即用酒精棉球擦拭迷宫中央区, 以消除气味影响。由于各组动物训练任务和处理不同, 所以训练天数也不尽相同。
1.2.3 悬尾应激方法 每天同一时间将悬尾组小鼠尾部悬挂于1.5 m高的木质横梁上持续20 min(图1C), 对照组小鼠仅接受相应的操作动作, 但未受实际的悬尾处理。观察悬尾处理对记忆获得的影响时, 悬尾后立即进行行为训练; 观察其对记忆巩固的影响时, 则每天在行为训练结束后立即进行悬尾;观察其对反转学习的获得或巩固时, 则分别在反转学习训练之前或之后进行悬尾。
1.3 数据统计分析
空间记忆作业正确次数的组间(悬尾处理动物与其对照组)比较均采用非配对样本t-检验(unpaired samplest-test)。动物训练成绩随训练时间的变化采用重复测量的双因子方差分析(two-way ANOVA with repeated measure)进行比较。所有比较均以P<0.05为差异显著标准。
2 结 果
2.1 悬尾处理小鼠的总体行为表现
动物在悬尾过程开始时, 挣扎激烈, 几分钟后,活动量减少,甚至不再有明显的肢体动作。悬尾过程中及之后, 多数小鼠排粪、排尿等应激反应明显增加。经连续5~7 d的20 min悬尾处理之后, 悬尾各组多数动物几乎不再出现明显的挣扎现象, 放回鼠笼后,其活跃程度明显低于对照组, 食量下降。
2.2 悬尾处理对小鼠绝对空间记忆的影响
在训练初期约1周左右(5~8 d)的时间内, 各悬尾组和其对照组的空间记忆成绩均处于随机水平(正确次数在3、4次左右), 未见明显差异(非配对样本t-检验:P> 0.05); 之后对照组动物的记忆成绩随训练时间的延长而显著提高, 悬尾组动物的成绩或呈略有提高或停滞不前, 与其对照组相比差异显著(非配对样本t-检验:P< 0.05; 重复测量的双因子方差分析:悬尾主效应,P< 0.01; 训练主效应,P<0.01; 悬尾×训练交互效应,P< 0.01)(图2, 图3)。以上数据表明, 悬尾处理可损伤小鼠绝对和相对空间记忆的获得和巩固过程。
2.3 悬尾处理对小鼠空间反转学习的影响
对照组动物在绝对和相对空间反转学习作业中的成绩经2~3 d训练后迅速提高, 悬尾组动物的成绩停滞不前。从第2~3 d开始, 各悬尾组与对照组的成绩出现显著差异(非配对样本t-检验:P<0.05; 重复测量的双因子方差分析:悬尾主效应,P<0.01; 训练主效应,P< 0.01; 悬尾×训练交互效应,P< 0.01)(图2, 图3)。以上数据表明, 悬尾处理可损伤小鼠绝对和相对空间反转学习的获得和巩固过程。
3 讨 论
3.1 悬尾处理损伤空间记忆及其反转学习的获得和巩固过程
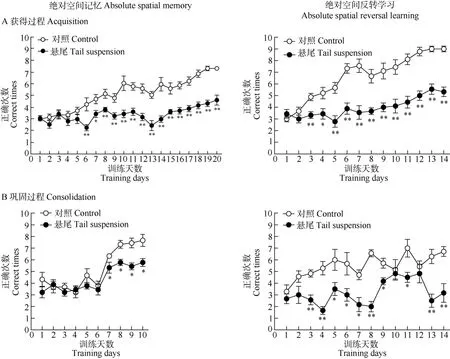
图2 悬尾处理对小鼠十字迷宫绝对空间记忆及其反转学习作业成绩的影响Fig. 2 The effects of tail suspension on the performance of absolute spatial memory and its reversal learning task in a plus maze in mice
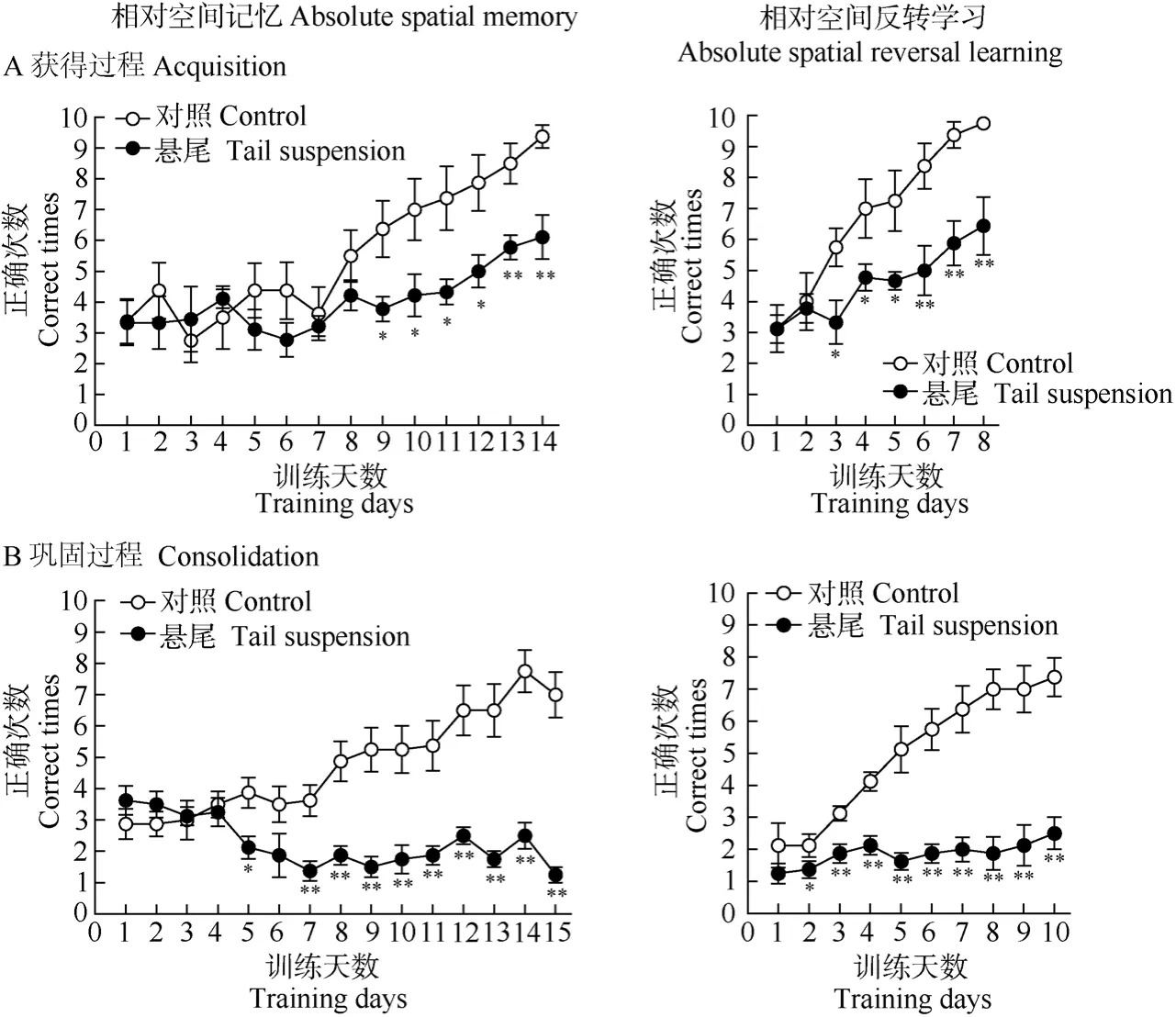
图3 悬尾处理对小鼠十字迷宫相对空间记忆及其反转学习作业成绩的影响Fig. 3 The effects of tail suspension on the performance of relative spatial memory and its reversal learning task in a plus maze in mice
本研究结果证明, 连续每天短时高空悬尾应激(20 min)可致小鼠绝对和相对空间记忆及其反转学习的获得和巩固过程显著受损, 其中相对空间记忆及其反转学习的巩固过程的受损更为严重。悬尾动物的成绩虽然明显低于对照组, 但随着训练也有所提高(在相对空间记忆及其反转学习的巩固过程中成绩未见明显提高), 说明悬尾动物的学习记忆功能处在相对缓慢的恢复过程中。在连续每天短时高空悬尾处理过程中, 动物可能会逐渐适应这种悬尾处理, 但能否完全恢复正常尚属未知。Wang et al (2005, 2007)采用一次性步入法发现使用同样的悬尾处理方法, 经单次高空悬尾处理20 min可导致被动回避行为, 即情绪记忆获得功能受损, 若悬尾处理后休息20 min后再测试, 则未见情绪记忆受损,说明单次悬尾的记忆损伤效应是暂时的、可逆的;但从本研究结果来看, 连续多天每天短时高空悬尾应激造成的记忆损伤效应可能因涉及的记忆过程、记忆信息类型不同而异。似乎相对空间记忆及其反转学习的巩固过程对悬尾处理更敏感一些。至于机体能否恢复正常, 取决于机体的适应调整能力能否消除损伤的累积效应。
本研究中一个值得注意的现象是, 在空间记忆训练初期约1周左右(5~8 d)的时间内, 各悬尾组和其对照组的空间记忆成绩均处于随机水平, 之后对照组动物成绩迅速提高, 两组之间拉开差距; 而在反转学习训练的第2~3 d开始, 对照组动物的成绩就已经显著高于悬尾组。最可能的一个原因是, 在训练初期, 对照组动物要适应训练规则, 并建立相应的行为反应策略和记忆的“空间认知地图”, 这可能需要较长的时间(大约1 周); 而在反转学习中,由于动物已在空间记忆训练中建立有效的反应策略, 反转学习无须改变策略, 只需要调整反应方向即可, 故可迅速作出调整。
3.2 悬尾处理与失重的记忆损伤效应
经典的头低位失重模型的应激作用较小, 主要用于探讨人类失重(下肢不负重)的长期效应 (Chen et al, 1993; Shen et al, 1989; Wu et al, 2000)。Wu et al (2000)使用矩形水迷津和跳台实验发现头低位悬尾模拟失重7 d或12 d可损伤小鼠绝对空间记忆、被动回避学习, 而悬尾5 h则未见损伤效应。另有报告指出, 头低位悬尾模拟失重7 d可造成大鼠分辨学习(Y-迷宫)和被动回避学习(避暗法)的明显损伤,这种损伤在模拟失重结束后经3 d的恢复仍未见明显改善 (Sun et al, 2009; Xu et al, 2005)。这些研究均提示, 长期失重可造成动物的学习和记忆损伤, 但短时间的头低位悬尾不会造成损伤。
以上模拟失重的方法与本研究采用的方法有所不同, 不同之处在于, 首先,前者采用大鼠或小鼠前肢着地、后肢离地,保持头部 ~ 30°下倾的悬尾倒立连续数天的方法(Chen et al, 1993); 而本研究和Wang et al (2005, 2007)采用的是头下尾上完全垂直的悬尾方法, 在更高的净水压力下, 会有更多体液向头胸部集中, 从而导致动物更强烈的应激。这也可能是本研究悬尾20 min即可致记忆损伤, 而Wu et al (2000)悬尾5 h未见记忆损伤的主要原因。其次, 本研究采用的是每天仅悬尾处理20 min, 而前者则是全天24 h一直给予悬尾处理。长时记忆主要分为三个阶段, 即获得、巩固和提取。前人的研究主要关注长期悬尾对学习和记忆的总体效应, 但不能说明其主要影响学习和记忆的哪个过程。而本研究则明确表明, 急性悬尾应激至少可损伤空间长时记忆及其反转学习的获得和巩固, 但是否影响已巩固记忆的提取过程尚需进一步探讨。
3.3 悬尾处理损伤记忆的可能机制
悬尾处理可引发机体生理和心理的各种急慢性应激反应。急性应激可通过HPA轴导致糖皮质激素释放增加, 并使交感神经兴奋, 脑内多种神经递质也会一过性的释放增加, 但GABA的释放减少,并涉及相应的受体 (Arnsten, 2009; Elzinga et al, 2005; Haggstrom et al, 1984; Mitsushima et al, 2008; Roozendaal et al, 2009; Tollenaar et al, 2008; Yuen et al, 2009)。慢性应激则可通过引发相关的基因表达以及神经元形态学的改变对包括学习和记忆在内的神经功能产生损伤作用(Lee et al, 2009; Mcewen, 2001; Mizoguchi et al, 2000)。以上应激效应也可在悬尾倒立模拟失重的动物或航天飞行动物上观察到(Cui et al, 2008; Sun et al, 2006, 2009; Zheng et al, 2008)。与其它应激方式不同, 悬尾处理可通过体位的转换, 使动物全身体液在重力的作用下发生头向再分配, 继而引发强烈的心血管反应, 并改变全身尤其是脑部血液循环、血液流变性以及血管、神经元超微结构的改变(Bi et al, 2004; Meng et al, 1997; Shen et al, 1989; Wu & Shen, 2000)。同时,体位的改变可干扰前庭等感觉器官、运动等系统的功能,使空间定向、运动知觉以及运动平衡、协调等能力受到损伤 (Shen, 2003; Yang & Shen, 2003)。除此之外, 在本研究中还观察到动物活动减少、食量降低的现象, 但不能确定此为悬尾应激的结果还是悬尾间接导致记忆损伤的原因之一。总的说来, 悬尾处理造成的影响与一般应激方式有相似之处, 但也有很多不同的地方, 是一种独特的应激方式。悬尾处理可能通过以上过程导致应激, 继而影响学习和记忆等认知功能。
致谢:云南师范大学生命科学学院2003级的汪和、王浩行和王伟花同学曾参与部分动物行为训练和数据收集等工作, 在此对他们的辛勤付出表示衷心的感谢!
Arnsten AF. 2009. Stress signalling pathways that impair prefrontal cortex structure and function [J]. Nat Rev Neurosci,10(6): 410-422.
Bi L, Li YX, He M, Huang ZM, Zhao DS, Lan JQ. 2004. Ultrastructural changes in cerebral cortex and cerebellar cortex of rats under simulated weightlessness [J]. Space Med Med Eng: Beijing,17(3): 180-183. [毕蕾,李英贤,何玫,黄增明,赵定胜,蓝景全. 2004.模拟失重对大鼠大脑和小脑皮层超微结构的影响.航天医学与医学工程,17(3): 180-183.]
Chen J, Ma J, Ding ZP, Zhang LF. 1993. A modified tail-suspension model for simulating long-term weightlessness [J]. Chn J Space Sci,13(2): 159-162. [陈杰,马进,丁兆平,张立藩. 1993.一种模拟长期失重影响的大鼠尾部悬吊模型.空间科学学报,13(2): 159-162.]
Cryan JF, Mombereau C, Vassout A. 2005. The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice [J]. Neurosci Biobehav Rev,29(4-5): 571-625.
Cui YP, Zhang F, Wang BC, Song YT, Yang WP, Zhu YL, Hong F. 2008. Comparative research of the effects of overload training and simulated weightlessness on the HPA axis in rats [J]. Chn J Sports Med,27(1): 90-92. [崔玉鹏,张凡,王宝成,宋玉涛,杨卫平,朱一力,洪峰. 2008.大负荷运动训练与模拟失重对大鼠HPA轴影响的对比研究.中国运动医学杂志,27(1): 90-92.]
Elzinga BM, Bakker A, Bremner JD. 2005. Stress-induced cortisol elevations are associated with impaired delayed, but not immediate recall [J]. Psychiatry Res,134(3): 211-223.
Haggstrom JE, Sjoquist B, Eckernas SA, Ingvast A, Gunne LM. 1984. Discrete regional distribution of biochemical markers for the dopamine, noradrenaline, serotonin, GABA and acetylcholine systems in the monkey brain (Cebus Apella). Effects of stress [J]. Acta Physiol Scand Suppl,534: 1-27.
Lee T, Jarome T, Li SJ, Kim JJ, Helmstetter FJ. 2009. Chronic stress selectively reduces hippocampal volume in rats: a longitudinal magnetic resonance imaging study [J]. Neuroreport,20(17): 1554-1558.
McEwen BS. 2001. Plasticity of the hippocampus: adaptation to chronic stress and allostatic load [J]. Ann N Y Acad Sci,933: 265-277.
Meng JR, Shen XY, Dong Q, Chen JH, Xiang QL, Wang DS. 1997. Effect of 7-day simulated weightlessness on hunman hemorheology [J]. J Chn Microcirc,1(2): 77-79. [孟京瑞,沈羡云,董颀,陈建和,向求鲁,汪德生. 1997. 7天模拟失重对人体血液流变性的影响.中国微循环,1(2): 77-79.]
Mitsushima D, Takase K, Funabashi T, Kimura F. 2008. Gonadal steroid hormones maintain the stress-induced acetylcholine release in the hippocampus: simultaneous measurements of the extracellular acetylcholine and serum corticosterone levels in the same subjects [J]. Endocrinology,149(2): 802-811.
Mizoguchi K, Yuzurihara M, Ishige A, Sasaki H, Chui DH, Tabira T. 2000.Chronic stress induces impairment of spatial working memory because of prefrontal dopaminergic dysfunction [J]. J Neurosci,20(4): 1568-1574.
Morey-Holton ER, Wronski TJ. 1981. Animal models for simulating weightlessness [J]. Physiologist,24(Suppl.): S45-46.
Morey-Holton ER, Globus RK. 2002. Hindlimb unloading rodent model: technical aspects [J]. J Appl Physiol,92(4): 1367-1377.
Roozendaal B, McReynolds JR, Van der Zee EA, Lee S, McGaugh JL, McIntyre CK. 2009. Glucocorticoid effects on memory consolidation depend on functional interactions between the medial prefrontal cortex and basolateral amygdala [J]. J Neurosci,29(45): 14299-14308.
Seong E, Seasholtz AF, Burmeister M. 2002. Mouse models for psychiatric disorders [J]. Trends Genet,18(12): 643-650.
Shen XY, Lan JQ, Xiang QL, Meng JR, Wang YQ. 1989. Changes of rabbit brain microcirculation during simulated weightless [J]. Space Med Med Eng: Beijing,2(2): 89-93, 151-152. [沈羡云,蓝景全,向求鲁,孟京瑞,王玉清. 1989.模拟失重时兔脑微循环的变化.航天医学与医学工程,2(2): 89-93, 151-152.]
Shen XY. 2003. Expectation of the study of weightlessness physiology in the 21st century [J]. Space Med Med Eng: Beijing,16(Suppl.): 573-576. [沈羡云. 2003. 21世纪失重生理学研究的展望.航天医学与医学工程,16(增刊): 573-576.]
Sun XQ, Xu ZP, Zhang S, Wu B, Cao XS, Wu P. 2006. Effects of +Gx exposure after simulated weightlessness on glial fibrillary acidic protein (GFAP) expression of brain in rats [J]. Space Med Med Eng: Beijing,19(1): 19-22. [孙喜庆,徐志鹏,张舒,吴斌,曹新生,吴萍. 2006.模拟失重后高+Gx作用对大鼠脑胶质细胞酸性蛋白表达的影响.航天医学与医学工程,19(1): 19-22.]
Sun XQ, Xu ZP, Zhang S, Cao XS, Liu TS. 2009. Simulated weightlessness aggravates hypergravity-induced impairment of learning and memory and neuronal apoptosis in rats [J]. Behav Brain Res,199(2): 197-202.
Tollenaar MS, Elzinga BM, Spinhoven P, Everaerd WA. 2008. The effects of cortisol increase on long-term memory retrieval during and after acute psychosocial stress [J]. Acta Psychol :Amst,127(3): 542-552.
Wang YF, Zhu B, Wu J, Wang L, Cai JX. 2005. The dopamine mechanism of impairment effect of tail-suspension on passive avoidance task in mice [J]. Neurosci Bull,21(Suppl.): 149. [王永富,朱壁,吴晶,王磊,蔡景霞. 2005.悬尾应激损伤小鼠被动回避作业的多巴胺受体机制.神经科学通报,21(增刊): 149.]
Wang YF, Cai JX, Xu L. 2007. I. Mechanism study on the anti-stress-like and anti-depressant-like effects of phenchlobenpyrrone, II. Synaptic plasticity mechanism of EGb 761- or Chinese-Tranditional-Medicine-Fufang on age-induced cognitive deficit in rats [PhD] [D]. Beijing: Graduate University of Chinese Academy of Sciences and Kunming Institute of Zoology, Chinese Academy of Sciences. [王永富,蔡景霞,徐林. 2007.Ⅰ、KMB-009改善心理应激所致认知障碍和抑郁样行为的研究 Ⅱ、银杏叶提取物及复方制剂改善老年鼠空间学习记忆突触可塑性机理[博士].北京:中国科学院研究生院 中国科学院昆明动物研究所.]
Wu DW, Shen XY. 2000. Changes of cerebral circulation during weightlessness or simulated weightlessness [J]. Space Med Med Eng: Beijing,13(5): 386-390. [吴大蔚,沈羡云. 2000.失重或模拟失重时脑循环的改变.航天医学与医学工程,13(5): 386-390.]
Wu DW, Shen XY, Dong Q, Wang SP, Cheng ZH, Zhang SJ. 2000. Effects of tail suspension on learning and memory function of mice [J]. Space Med Med Eng: Beijing,13(4): 244-248. [吴大蔚,沈羡云,董颀,王圣平,程张红,张淑静. 2000.尾部悬吊对小鼠学习记忆功能的影响.航天医学与医学工程,13(4): 244-248.]
Xu ZP, Sun XQ, Liu TS, Wu B, Zhang S, Wu P. 2005. Effects of high +Gx during simulated spaceship emergency return on learning and memory in rats [J]. Space Med Med Eng:Beijing,18(1): 1-5. [徐志鹏,孙喜庆,刘挺松,吴斌,张舒,吴萍. 2005.模拟飞船应急返回时高+Gx对大鼠学习记忆功能的影响.航天医学与医学工程,18(1): 1-5.]
Yang JJ, Shen Z. 2003. Effects of microgravity on human cognitive function in space flight [J]. Space Med Med Eng :Beijing,16(6): 463-467. [杨炯炯,沈政. 2003.载人航天中微重力环境对认知功能的影响.航天医学与医学工程,16(6): 463-467.]
Yuen EY, Liu W, Karatsoreos IN, Feng J, McEwen BS, Yan Z. 2009. Acute stress enhances glutamatergic transmission in prefrontal cortex and facilitates working memory [J]. Proc Natl Acad Sci U S A,106(33): 14075-14079.
Zheng LJ, Liu L, Zhang T, Lü BC. 2008. The study of the hippocampal dentate gyrus neurogenesis after simulated weightlessness in the rat [J]. Chn J Neuroanat,24(3): 287-291. [郑丽娟,刘立,张婷,吕伯昌. 2008.模拟失重后大鼠海马齿状回神经发生的实验研究.神经解剖学杂志,24(3): 287-291.]
Impairment effects of tail suspension stress on spatial memory and its reversal learning in mice
WANG Xiao-Qin1, WANG Gong-Wu2,3,*, DUAN Qiu-Yan2, TENG Shi-Tong2
(1. School of Physical Education, Yunnan Normal University, Kunming 650500, China; 2. Laboratory of Behavioral and Cognitive Neurobiology, School of Life Sciences, Yunnan Normal University, Kunming 650500, China; 3. Engineering Research Center of Sustainable Development and Utilization of Biomass, Ministry of Education & Key Laboratory of Yunnan for Biomass Energy and Biotechnology of Environment, Kunming 650500, China)
Present work investigated the effects of tail suspension stress (TSS) on spatial memory acquisition, consolidation, and its reversal learning in mice. Eighty-one adult male KM mice were divided into four groups (each group including a TSS subgroup and its control subgroup): absolute spatial memory acquisition and consolidation groups (group AA and CA); relative spatial memory acquisition and consolidation groups (group AR and CR). TSS (20 min) was performed immediately before (acquisition) or after (consolidation) a daily training. Results showed that there was no significant difference between control animals and TSS animals in each group in early spatial memory training days (5-8 d of training). Along with training, the performance of control animals improved significantly, but the performance of TSS animals improved slightly (group AA, CA and AR) or even did not change (group CR) (P< 0.01). Reversal learning was also impaired in TSS animals (P< 0.01). The results indicated that TSS could impair spatial memory acquisition, consolidation and reversal learning (especially the relative spatial memory consolidation and its reversal learning) in mice.
Tail suspension stress; Spatial memory; Reversal learning; Acquisition; Consolidation; KM mice
Q189; Q427
A
0254-5853-(2011)04-0428-07
10.3724/SP.J.1141.2011.04428
2011-06-05;接受日期:2011-06-28
中国科学院“西部之光”人才培养计划项目(2008);云南省运动人体科学实验教学示范中心建设项目(2009);国家自然科学基金项目(31060142)
∗通讯作者(Corresponding author),王功伍, Tel: +86-871-5516068, Fax: +86-871-5516759, E-mail: waov@tom.com
王晓琴(1978-),硕士,讲师,主要从事运动生理学和神经生物学研究;E-mail: xiaoqinwang2001@yahoo.com.cn
猜你喜欢
杂志排行

Zoological Research的其它文章
- Metabolism and thermoregulation between Mrs Hume’s Pheasant (Syrmaticus humiae) and Elliot’s Pheasant (S. ellioti)
- Behavioral migration diversity of the Yangtze River Japanese Eel, Anguilla japonica, based on otolith Sr/Ca ratios
- Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth’s Leaf Warbler (Phylloscopus reguloides)
- Afferent and efferent pathways in the visual system of the freshwater snail Planorbarius corneus
- Notch signaling dependent differentiation of cholangiocyte-like cells from rhesus monkey embryonic stem cells
- Localization of stationary pronuclei during conjugation of Paramecium as indicated by immunofluorescence staining