野桑蚕丝胶1基因Ser1A′及其上游调控序列的克隆和序列分析*
2011-06-02陈典刚张永红李春峰周泽扬
黄 科 伍 杰 陈典刚 张永红 李春峰 周泽扬,3
(1.西南大学蚕学与系统生物学研究所,生物技术学院,重庆 400716;2.重庆文理学院花卉研究所,重庆 402160;3.重庆师范大学生命科学学院,重庆 401331)
蚕丝蛋白主要由丝素和丝胶组成。丝胶蛋白在中部丝腺特异表达,主要由丝胶1蛋白和丝胶2蛋白组成[1-2]。家蚕丝胶1蛋白由4条不同的mRNA编码,由同一条mRNA经不同的剪切方式形成,其长度分别为10.5kb、9.0kb、4.0kb和2.8kb[3]。1997年Garel A等比较系统地阐明了家蚕4.0kb丝胶1基因的mRNA序列,在家蚕基因组中丝胶1基因含8个内含子[4]。丝胶2基因也通过不同的剪切方式形成两条mRNA编码丝胶2蛋白[2]。丝胶1蛋白和丝胶2蛋白的表达受到组织的严格调控,也受到幼虫发育时期的调控[5]。基因转录调控中,启动子是关键,启动子序列所含的顺式作用元件与转录调控因子的结合决定基因的转录。家蚕丝胶1基因启动子中存在三个顺式作用元件SA(-103~-85)、SB(-149~-135)和SC(-204~-183),都能特异的与转录因子(SGF-1,SGF-3)结合[6]。丝腺转录因子1(SGF-1)属于head/HNF-3家族,能特异结合SA位点,认为决定了丝胶1基因的组织特异性表达[7]。体外转录实验表明去除SA元件能下调丝胶1基因启动子的活性[6]。丝腺转录因子3(SGF-3)在丝腺核抽提物中含量最多,能与SC位点特异的结合,体外转录实验同样证明去除SC位点丝胶1基因启动子的活性也存在下调[8]。
我们在克隆家蚕丝胶1基因上游调控序列构建中部丝腺特异表达转基因载体时,发现不同家蚕品系丝胶1基因上游调控序列存在差异,在调查的66个家蚕品系中,丝胶1基因上游调控序列的-1015~+48位置存在多态性,PCR扩增结果有1063bp、636bp及同时出现这两种条带的情况[9]。家蚕(Bombyx mori)是由野桑蚕(Bombyx mandarina)驯化而来,二者的亲缘关系很近。家蚕经历了强烈的人工选择,成为了完全驯养的昆虫,并依赖人类而生存,而野桑蚕完全在自然条件下生存。与家蚕相比,野桑蚕具有较强的疾病防御及抗逆能力,但其蚕茧大小、生长速率等经济性状则显著弱于家蚕。本实验调查了不同地方的野桑蚕丝胶1基因上游调控序列,结果表明野桑蚕 sericin1上游调控序列也存在多态性,分别为636bp(HQ702378)及1061bp(HQ702379)。对野桑蚕丝胶1基因(HQ702380)编码序列进行克隆,通过序列比对,分析家蚕与野桑蚕丝胶1基因同源序列之间的差异,旨在为进一步了解丝胶1基因的表达调控有所帮助。
1 材料与方法
1.1 材料与试剂
野桑蚕材料来自山东、重庆青木关、四川泸州、资阳、南充、湖南、苏州和日本。克隆用宿主菌DH5α、质粒pSLfa1180fa由本实验室保存。DNA聚合酶、M-MLV反转录酶购自PROMEGA 公司 ;限制性内切酶 Hind III、Eco RI、Mlu I、Sal I和 Sma I及 pMD18-T-vector购于TaKaRa公司;胶回收试剂盒购于上海华舜公司。
1.2 野桑蚕丝胶1基因的克隆
以GenBank中登录的家蚕丝胶1基因(sericin 1A)的核酸序列(AB112019.1)设计引物,Ser1 F:5'ATGCGT TTCGT TCTGTGCTGC 3',Ser1 R:5'T TAGT TGTATTAAACACCGAT 3',扩增野桑蚕丝胶1基因。参照T ripure试剂说明提取重庆青木关野桑蚕5龄第3天丝腺总RNA,然后以野桑蚕总RNA为模板,按M-MLV反转录酶的使用说明进行cDNA第一链的合成。以野桑蚕cDNA为模板,用引物对Ser1 F/Ser1 R进行PCR扩增,反应条件为:94 ℃预变性4 min;94℃30 s,55℃40 s,72 ℃4 min,35个循环;72 ℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳,切胶回收目的产物,克隆到pMD18-T-vector中,酶切鉴定后送交上海英俊公司进行测序。
1.3 野桑蚕丝胶1基因上游调控序列的克隆
根据GenBank中登录的家蚕丝胶1基因上游调控序列(AB007831.1)和家蚕丝胶1基因mRNA序列(AB112019.1)设计引物,上游引物为 Ser1p F:5'AAAGCATAAGCGGTCAGAAACC 3',下游引物 Ser1p R:5'GGTCTT TGGATCGCTTGATCC 3',通过PCR扩增野桑蚕丝胶1基因上游调控序列。野桑蚕基因组DNA的抽提采用蛋白酶K/RNA酶A的方法提取。以抽提的野桑蚕基因组DNA为模板,用引物对Ser1p F/Ser1p R进行PCR扩增,反应条件为:94℃预变性4min;94℃50 s,60℃55 s,72℃90 s,35个循环;72℃延伸 10min。将PCR产物回收、克隆、酶切鉴定并测序。
1.4 序列分析
克隆的野桑蚕丝胶1基因序列用DNAStar软件预测其蛋白质序列与家蚕Sericin 1 A′(Ser 1A′,BAD00699.1),sericin 1A(Ser 1A ,BAD00698.1),sericin 1B(Ser 1B ,BAD00700.1),sericin 1B(ser 1B,Q17240)作muscle序列分析。将测定的野桑蚕丝胶1基因上游调控序列和家蚕品系J115 Sericin 1上游调控序列及GenBank中登录的Sericin 1上游调控序列AB007831.1作muscle序列分析。
2 结果
2.1 野桑蚕丝胶1基因的克隆
以5龄3天野桑蚕丝腺cDNA为模板,用引物对Ser1 F/Ser1 R扩增得到一条约2.1 kb的条带。将该片段回收连接到pMD18-T载体中,用Hind III和EcoRI双酶切鉴定筛选的重组质粒,酶切出一条长度约2.1 kb的片段(图1)。阳性克隆进行测序后,由于野桑蚕丝胶1基因重复序列较多,导致不能拼接出完整的ORF序列。针对此种情况,先将克隆的野桑蚕丝胶1基因酶切,形成较小的片段后进行亚克隆,亚克隆测序后再进行拼接。将克隆的野桑蚕丝胶1基因用多种限制性内切酶分别进行酶切,结果发现Mlu I可以将克隆的野桑蚕丝胶1基因酶切成约700 bp和1.4 kb的两个片段。利用pMD18-T vector上Sal I和Sma I酶切位点,将克隆的野桑蚕丝胶1基因同时以 Mlu I、Sal I和Sma I进行酶切,回收700 bp小片段亚克隆到pSLfa1180fa载体的Mlu I、Sal I位点,回收1.4 kb大片段亚克隆到pSLfa1180fa载体的Mlu I、Sma I位点。将两个亚克隆经酶切鉴定(图2)及测序拼接,得到一条完整的ORF序列(GenBank登录序列号:HQ702380)。
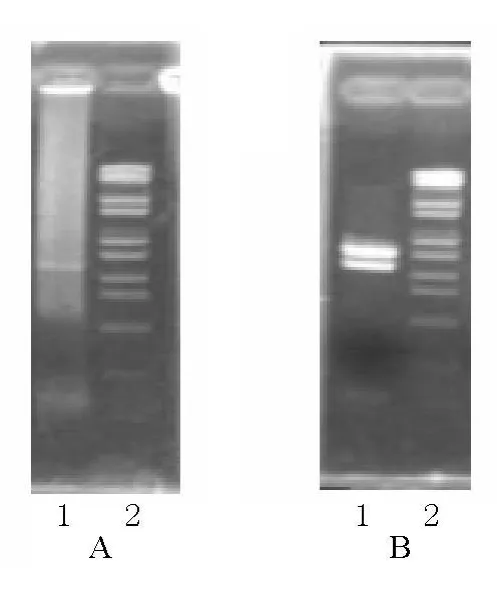
图1 野桑蚕丝胶1基因的PCR及重组质粒的酶切检测

图2 亚克隆酶切鉴定
2.2 野桑蚕丝胶1基因上游调控序列克隆
以8个不同地方野桑蚕基因组DNA为模板,用引物对Ser1p F/Ser1p R进行野桑蚕丝胶1基因上游调控序列PCR扩增,从山东野桑蚕基因组DNA中扩增出一条约1kb条带,从湖南野桑蚕、四川南充野桑蚕和重庆青木关野桑蚕基因组DNA中扩增出一条约600bp条带,另外4个样品没有扩增结果(图3)。将扩增出约600bp条带的青木关野桑蚕及扩增出1kb条带的山东野桑蚕的PCR产物回收后,克隆到pMD18-T载体。重组质粒经过酶切验证后进行测序。
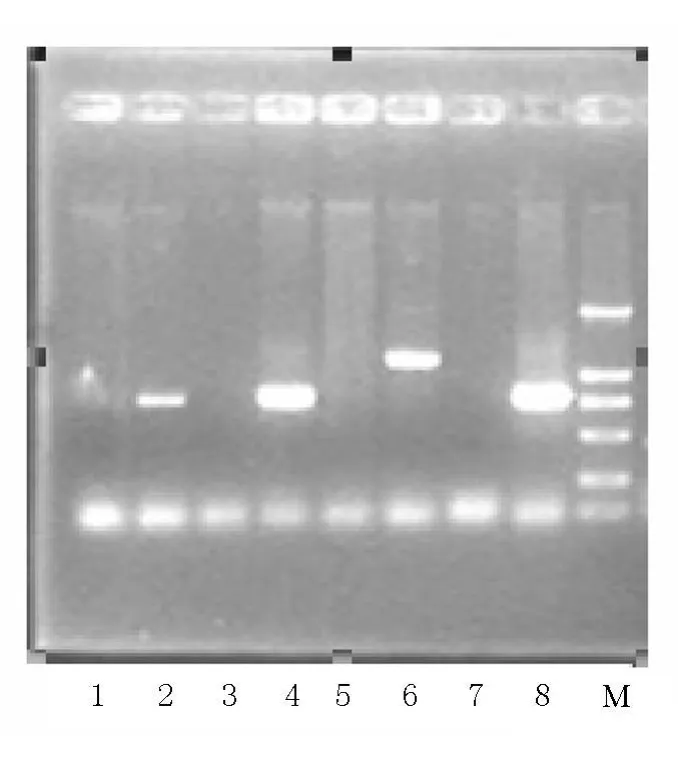
图3 野桑蚕Sericin1启动子PCR扩增
2.3 野桑蚕丝胶1基因序列分析
克隆的野桑蚕Sericin 1序列用DNAStar预测其蛋白质序列与家蚕Sericin 1A′(Ser1 A′,BAD00699.1)、Sericin 1A(Ser1 A ,BAD00698.1)、Sericin 1B作 muscle序列分析(图4),结果表明:预测的野桑蚕丝胶1蛋白与家蚕Sericin 1 A′、Sericin 1A、Sericin 1B(Ser 1B,BAD00700.1)、Sericin 1B(ser 1B,Q17240)有较强的相似性,相似性分别为 98%、98%、92%、97%。据Garel等预测的丝胶1蛋白(Ser 1B,Q17240)一级结构中,Ser 1B多肽中含1、2、3、4、5、7、8、9外显子[4],而野桑蚕丝胶 1蛋白与家蚕Ser1 A′相似性最高,其只含1、2、7、8、9外显子,因此将克隆的2176bp野桑蚕丝胶1基因命名为B.mandarin Sericin1 A′。野桑蚕丝胶蛋白中存在由38个氨基酸组成的重复序列,包含约40%的丝氨酸(图5),正是由于这些重复序列导致测序后拼接非常困难,利用内切酶切成小片段亚克隆后才完成拼接。
2.4 野桑蚕丝胶1基因上游调控序列分析
将克隆的野桑蚕Sericin 1上游调控序列与我们之前克隆的家蚕J115品系Sericin 1上游调控序列,及GenBank中登录的Sericin 1上游调控序列AB007831.1作muscle序列分析,结果表明克隆的山东野桑蚕Sericin 1上游调控序列、青木关野桑蚕Sericin 1上游调控序列片段和家蚕Sericin 1上游调控序列具有很强的相似性。1061bp丝胶1基因上游调控序列是636bp丝胶1基因上游调控序列中插入约400bp的片段,而前后约300bp是一致的,克隆的野桑蚕丝胶1基因上游调控序列中也存在SA、SB和SC基序(图6)。克隆的野桑蚕丝胶1基因上游调控序列在GenBank中登录序列号分别为HQ702380和HQ702379。
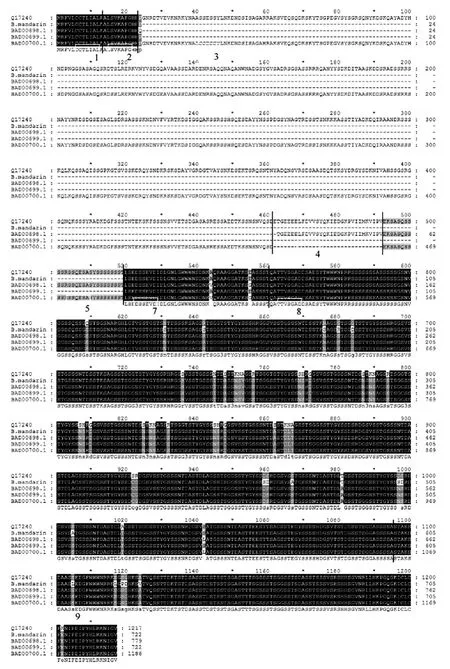
图4 预测的野桑蚕丝胶1蛋白序列与家蚕丝胶1蛋白序列比对

图5 野桑蚕丝胶1蛋白序列的重复基序

图6 野桑蚕Sericin 1上游调控序列与家蚕Sericin 1上游调控序列比对分析
3 讨论
家蚕丝胶1基因在基因组中是单拷贝的,转录后通过不同的剪切方式形成4条mRNA,其长度分别为10.5kb、9.0kb、4.0kb和2.8kb[3]。Garel等指出在家蚕丝胶1基因剪切时1、2、7、8和9外显子存在于所有形式的mRNA中,而3、4、5和6外显子是选择性剪切的,所有外显子都包含的丝胶1基因mRNA命名为Ser1C,不包含3和4外显子的丝胶1基因mRNA命名为Ser1D;不包含外显子6的丝胶1基因mRNA命名为Ser1B;不包含外显子3和6的丝胶1基因mRNA命名为Ser1A;不包含外显子3、4、5和6的丝胶 1基因mRNA命名为Ser1A′[4]。而本实验克隆的重庆野桑蚕丝胶1基因编码序列与家蚕Ser1A′相似性非常高,其仅包含丝胶1基因的1、2、7、8和9外显子,故将其命名为野桑蚕Ser1A′。野桑蚕Ser1A′富含丝胶1基因的特征性重复基序,正是由于重复基序的存在,完整的野桑蚕Ser1A′编码序列在测序时形成特殊的结构而造成测序困难,而将野桑蚕Ser1A′编码序列酶切为较小的两个片段后,分别克隆到pSLfa1180载体然后进行测序,由于序列重复度减小,两个亚克隆测序后经拼接得到完整的Ser1A′编码序列。
在调查不同地方野桑蚕丝胶1基因上游调控序列多态性时,发现与家蚕相似,也存在1061bp与636bp两种PCR扩增情况,但没有发现在同一个地方个体中同时存在636bp和1061bp的情况,而这种情形在家蚕中也是存在的[9],这可能与我们调查的同一个地方样品数量少有关。除此之外,野桑蚕丝胶1基因上游调控序列与家蚕丝胶1基因上游调控序列间存在极强的相似性,丝胶基因表达调控的三个顺式作用元件都位于相应的位点。
家蚕由野桑蚕驯化而来,目前对家蚕丝胶基因的研究比较多,而对野桑蚕丝胶1基因的研究报道较少,本研究在家蚕丝胶1基因研究基础上,仅克隆了野桑蚕丝胶1基因Ser1A′,而野桑蚕中的丝胶1基因其它不同剪切方式则有待研究。
[1]Okamoto H ,Ishikawa E ,Suzuki Y.Structural analysis of sericin genes.Homologies with fibroin gene in the 5'flanking nucleotide sequences[J].J Biol Chem ,1982,257(24):15192-15199.
[2]Michaille J J,Garel A ,Prudhomme J C.Cloning and characterization of the highly polymorphic Ser2 gene of Bombyx mori[J].Gene,1990 ,86(2):177-184.
[3]Michaille J J,Couble P ,Prudhomme J C,et al.A single gene produces multiple sericin messenger RNAs in the silk gland of Bombyx mori[J].Biochimie,1986,68(10-11):1165-1173.
[4]Garel A ,Deleage G ,Prudhomme J C.Structure and organization of the Bombyx mori sericin 1 gene and of the sericins 1 deduced from the sequence of the Ser 1B cDNA[J].Insect Biochem Mol Biol,1997 ,27(5):469-477.
[5]Couble P,Michaille J J,Garel A,et al.Developmental switches of sericin mRNA splicing in individual cells of Bombyx mori silkgland[J].Dev Biol,1987,124(2):431-440.
[6]M atsuno K ,Hui C C,Takiya S,et al.Transcription signals and protein binding sites for sericin gene transcription in vitro[J].J Biol Chem,1989,264(31):18707-18713.
[7]M ach V ,Takiya S ,Ohno K ,et al.Silk gland factor-1 involved in the regulation of Bombyx sericin-1 gene contains fork head motif[J].J Biol Chem ,1995 ,270(16):9340-9346.
[8]M atsuno K ,Takiya S ,Hui C C ,et al.T ranscriptional stimulation via SC site of Bombyx sericin-1 gene through an interaction with a DNA binding protein SGF-3[J].Nucleic Acids Res,1990 ,18(7):1853-1858.
[9]黄科,李春峰,周泽扬.Seriein 1上游调控序列多态性分析[J].西南师范大学学报(自然科 学版),2006,31(5):154-158.
