3种彩色叶玉簪光合日变化1)
2011-05-31李金鹏
李金鹏 董 然
(吉林农业大学,长春,130118)
玉簪属(Hosta)为百合科(Liliaceae)多年生草本植物,主要分布在东亚的温带与亚热带地区,即日本、朝鲜和中国的东部和南部,其中中国原产4个种,目前栽培品种已在世界各地广为栽培与应用,尤其近年一些花叶或彩叶品种在国外育种和应用十分普遍,现世界已命名的玉簪属园艺栽培品种达4 000个以上[1]。该属植物是中国的传统花卉,其株型、叶色、叶型丰富且花型美丽,国内部分品种已应用于园林造景和室内观赏。
本研究以金头饰玉簪(H.‘Golden Tiara’)为试材,其叶片心形,中部深绿,边缘为不规则的淡黄色或黄绿色,交界处条状交叉,侧脉6对;金鹰玉簪(H.‘Gold Edger’)叶片黄绿色、有光泽,卵形或心卵形,先端渐尖,叶脉深陷具7~9对侧脉;黄金叶玉簪(H.Sieboldiana‘Gold’)叶色浅黄绿至黄色,卵形或心卵形,具4~5对侧脉且叶片较薄。上述3种玉簪均是从国外引进的植株低矮丛生型彩色叶品种,引种发现其繁殖速度快,花葶均高出叶片,花色淡紫、漏斗状,花、叶观赏价值高,是难得的春夏观叶赏花的地被花卉。目前,国内外许多学者对玉簪属植物的研究主要集中在其系统分类、育种、组织培养、栽培及药理等方面[2-4],而对其光合生理和蒸腾特性方面的研究较少,尤其缺乏不同叶色品种的对比研究。金头饰、金鹰与黄金叶玉簪分别是花叶、黄绿叶、黄叶类型的玉簪品种,关于这3种彩叶玉簪品种的光合日变化对比研究未见报道。
本文从光合生理研究入手,探讨在自然光照条件下3种彩叶玉簪的光合特性,比较了不同叶色品种净光合速率的差异,分析其净光合速率日变化与主要生理生态因子的关系,并根据光补偿点和光饱和点判断其耐荫性,以期为3种玉簪的引种驯化、栽培应用提供理论依据。
1 材料与方法
试验地位于长春市吉林农业大学园林基地,地处湿润区向亚干旱区的过渡地带,属大陆性季风气候,其春季干燥多风,夏季温热多雨,秋季晴朗温差大,冬季漫长较寒冷。年平均气温4.8℃,夏季7月份最高温度39.5℃,冬季1月份最低温度-39.8℃,年平均降水量为567.0 mm,夏季降水量占全年降水量的60%以上,是降水量最丰富时期。
供试材料:金头饰玉簪(H.‘Golden Tiara’)、金鹰玉簪(H.‘Gold Edger’)及黄金叶玉簪(H.sieboldiana‘Gold’)。2007年4月从中国科学院北京植物所引入,定植在园林基地,经3 a引种栽培,表现良好,于2010年5月中旬将分株苗移植到20 cm×20 cm的黑色塑料盆钵中,在50%遮荫棚缓苗30 d后移置自然光照条件下培养,给予正常肥水管理。
供试土壤性质:pH值6.2、有机质质量分数7.68 g·kg-1、碱解氮质量分数 138.24 mg·kg-1、速效磷质量分数 42.37 mg·kg-1、速效钾质量分数 262.34 mg·kg-1。
光合作用日变化测定:利用美国PPSystems公司生产的CIRAS-2便携式光合仪进行测定。选择植株生长状况一致的向阳叶片(每个品种3株,每株选择1片叶,每叶测3次,共9个重复),于2010年7月中旬晴朗天气6:00—18:00每隔2 h测定1次。测定叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(Ci),光合有效辐射(PAR)、外界CO2摩尔分数以及气温、相对湿度(RH)等相关指标。
光响应曲线测定:于2010年7月中旬晴朗天气测定PAR在0~ 1400μmol·m-2·s-1范围内的Pn值,设定梯度为1 400、1 000、800、600、400、200、150、100、50、0 μmol·m-2·s-1。Pn对光照强度的响应进程,用叶子飘模型进行拟合[5],该模型的表达式为:Pn=a(1-βI)(I-LCP)/(1+γI),其中 Pn是净光合速率,α是光响应曲线的初始斜率,β称为修正系数,γ=α/Pmax(Pmax是植物的最大净光合速率),I为光照强度,LCP光补偿点。绘制光合—光响应曲线并分别求出光补偿点(LCP)、光饱和点(LSP)、表观量子效率;叶片水分利用效率用Pn与Tr的比值来表示。采用Excel2003、SPSS11.5分析软件进行数据处理,用SigmaPlot10.0进行作图。
2 结果与分析
2.1 光合生理参数日变化情况
光合作用日变化是植物生产过程中物质积累与生理代谢的基本单元,常因植物种类和环境条件的变化而有差异,它是分析环境因素影响植物生长和代谢的重要手段[6]。3种彩叶玉簪光合参数见表1。
由表1可知,在自然条件下3种玉簪的净光合速率、气孔导度、蒸腾速率的日变化趋势大致相似,均呈现“双峰”曲线,2次高峰出现的时间分别为10:00和16:00,仅各别品种的指标高峰出现时间有所差异,谷值出现在14:00左右,表现出典型的“午休”现象。其中3种玉簪的净光合速率第2峰值明显低于第1峰值,均是对午前上升的光照强度比较敏感,随着光照强度增强,净光合速率开始增加,表明均能充分利用上午的有利条件进行光合作用,1 d内净光合速率的大小依次为:金头饰玉簪>金鹰玉簪>黄金叶玉簪;金鹰玉簪的气孔导度在第1峰值发生时刻与其它两种玉簪有所差异,约在8:00左右出现,表明8:00左右的光照强度有利于金鹰玉簪气孔开张,气孔阻力降低,气孔导度增大;并且金鹰玉簪在8:00左右蒸腾速率达到1 d中的最高峰值,这一结果可能与气孔的调节有关[7],金头饰、黄金叶玉簪的变化与金鹰玉簪差异相对较大,出现在10:00,且在14:00以后没有下降的现象,由此可以看出不同叶色之间蒸腾速率差异较大。
3种玉簪的胞间CO2摩尔分数在1 d中均呈早晚高、中午又回升的变化趋势,近“W形”,中午胞间CO2摩尔分数回升可能是由于非气孔因素,阻碍了CO2的利用,致使植物体内细胞间隙的CO2增加,但3种玉簪中午胞间CO2摩尔分数并未超过早晚值,说明这两种玉簪中午利用CO2能力高于早晚。早6:00时3种玉簪胞间CO2摩尔分数范围在364~390 μmol·mol-1,随着 Pn的加强,出现“午休”现象,第 1 个低谷值出现在10:00左右(金头饰玉簪、金鹰玉簪)或12:00左右(黄金叶玉簪),第2个低谷值出现在16:00左右。
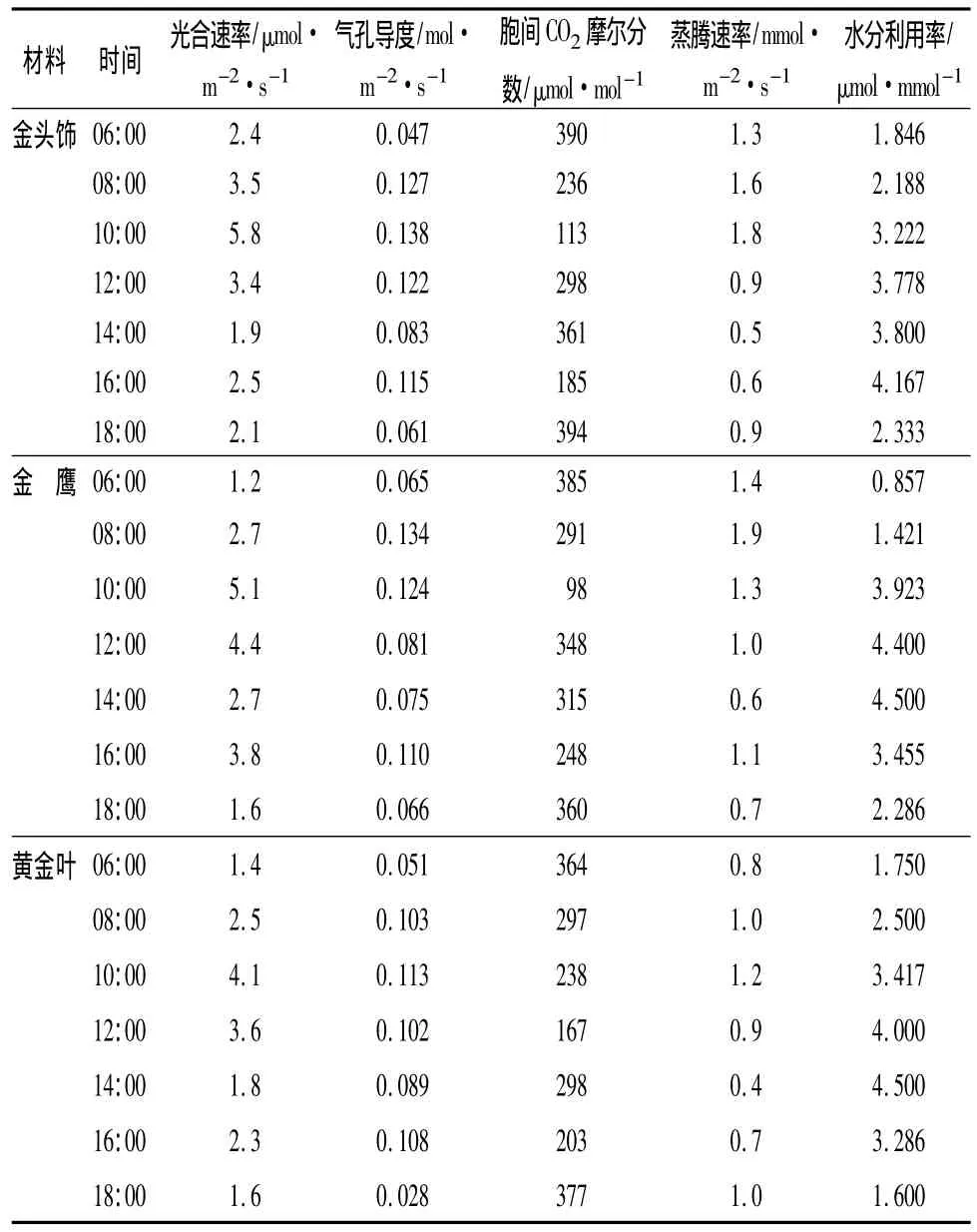
表1 3种玉簪光合作用日变化指标比较
水分利用效率是植物吸收利用水分的一个非常重要的指标,在1 d中会受环境因子直接或间接的影响而不断变化。表1所示,3种玉簪水分利用效率均呈现单峰曲线,在14:00左右(金鹰玉簪和黄金叶玉簪)或16:00左右(金头饰玉簪)出现高峰,3种玉簪材料的日平均水分利用效率从高到低排序为金头饰玉簪>黄金叶玉簪>金鹰玉簪。由此可见,不同叶色的玉簪,其水分利用效率是不同的。花叶的金头饰玉簪具有较强的水分利用能力,表现抗旱能力相对较强,而黄叶的黄金叶与黄绿叶的金鹰玉簪次之。
2.2 净光合速率与其他影响因子的关系
光合作用常受到植物内部生理状态和外界环境因子的共同制约,以其测定时刻的光合特征参数平均值进行相关性分析(净光合速率与其影响因子光合有效辐射、胞间CO2摩尔分数、气孔导度、蒸腾速率、气温、外界CO2摩尔分数、相对湿度),结果见表2。3种玉簪材料的净光合速率与胞间CO2摩尔分数呈显著性负相关(P<0.05),金头饰玉簪的净光合速率与蒸腾速率呈显著正相关(r=0.780*,P<0.05),金鹰玉簪与黄金叶玉簪净光合速率均与光合有效辐射呈显著正相关,与其他因子呈不显著正或负相关关系。从相关系数的大小来看,金头饰玉簪的蒸腾速率相关系数最大,即其蒸腾速率对净光合速率的影响最为明显,其次是胞间CO2摩尔分数;其余两种供试玉簪光合有效辐射相关系数最大,即这两种玉簪的净光合速率最大影响因子是光合有效辐射,其次是胞间CO2摩尔分数,这可能与光合有效辐射的变化幅度大有关系。

表2 净光合速率与各因子的相关性分析
2.3 3种玉簪的光响应曲线参数
光响应曲线可反映植物净光合速率随光合有效辐射增减的变化规律。表3所示:3种玉簪光响应曲线变化规律相似,当光合有效辐射为0 μmol·m-2·s-1时,净光合速率为负值,随着光合有效辐射的增强,净光合速率逐渐增加由负值转为正值,当净光合速率为0时的光合有效辐射即为光补偿点,当光合有效辐射在800 μmol·m-2·s-1时净光合速率达到最大值,该点即为光饱和点(其中金头饰玉簪在600 μmol·m-2·s-1);当光合有效辐射超过一定的范围时,净光合速率稍微略有下降的趋势,光合有效辐射过强会造成光合速率下降或者出现光抑制现象,表明3种玉簪对强光的适应能力较弱。
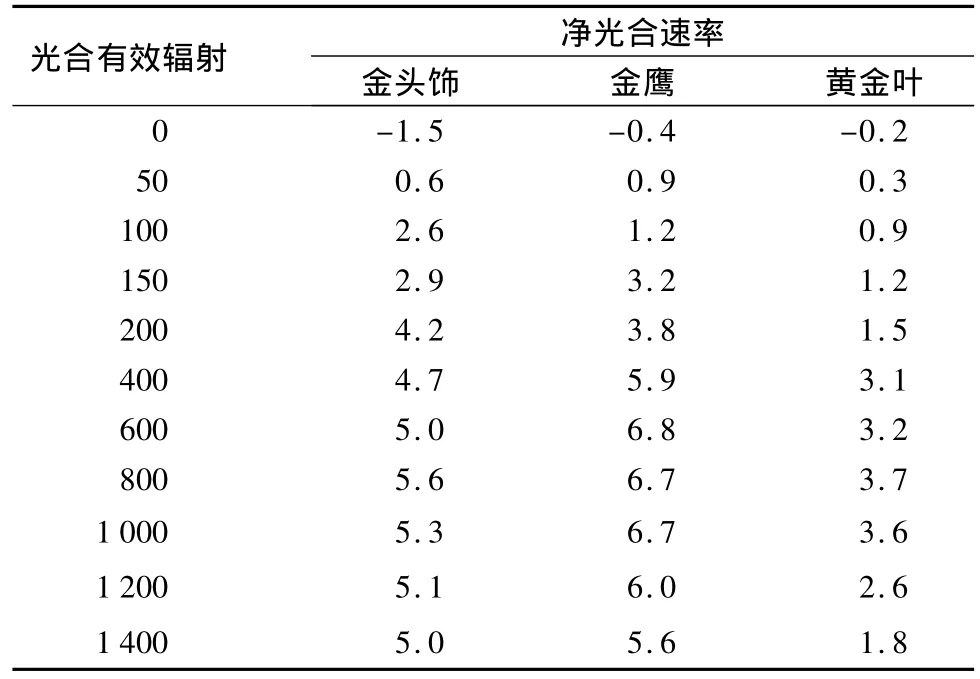
表3 3种玉簪的净光合速率与光合有效辐射变化规律μmol·m-2·s-1
从表4可知,3种玉簪的光补偿点、光饱和点与表观量子效率和Pmax差异显著,不同叶色玉簪净光合速率对光响应的结果有明显的差异。根据光响曲线的模型计算出3种玉簪叶片的光补偿点、光饱和点和表观量子效率和最大净光合速率等光合参数,其中金头饰玉簪的光饱和点最大(802.58 μmol·m-2·s-1),而光补偿点最小(17.493 μmol·m-2·s-1),表明花叶的金头饰玉簪对光环境的适应性较宽,不仅能充分利用弱光,而且对强光的承受能力也相对较强,其他两种叶色的玉簪则相对较弱,其中黄绿叶的金鹰玉簪耐荫性最弱。

表4 3种玉簪的光响应曲线参数
3 结论与讨论
植物在自然条件下,其光合作用日变化的曲线大致有3种类型,一种是中午光合速率最高的单峰曲线,另一种是在光照强度和水分亏缺的情况出现明显的光合“午休”现象,即日变化表现上、下午各有一个高峰的双峰曲线[8-9]和三峰曲线[10]。柯世省[11]认为“午休”现象是植物在高温强光条件下出现的普遍现象。林夏珍[12]等测定窄头橐吾(Ligularia stenocephala)光合特性日变化,发现该种植物净光合速率日变化是双峰型,本研究与其结果一致。3种玉簪的净光合速率日变化呈典型的“双峰”曲线,峰值分别出现在10:00和16:00,中午出现低谷,出现了光合“午休”现象,总体看金头饰玉簪净光合速率最高,表现出较高的光合效率,而黄金叶玉簪最低,金鹰玉簪介于二者之间,可见花叶的金头饰玉簪光能利用率高于其他两种叶色玉簪。此外,研究发现3种玉簪光合作用的主要时期是属于上午类型,而有些草坪草则下午保持较高水平的光合效率[13]。
陶汉之[14]和许大全等[15]认为气孔导度对净光合速率的影响并不明显,本研究结论:3种玉簪净光合速率与影响因子胞间CO2摩尔分数呈显著负相关,金头饰玉簪的净光合速率与蒸腾速率呈显著正相关,金鹰与黄金叶玉簪净光合速率均与光合有效辐射呈显著正相关,与其他因子呈不显著正或负相关关系,表明与前人观点一致。许多研究认为[16-17],叶片净光合速率与自身因素和环境因子密切相关,影响净光合速率的因素中,既有气孔限制又有非气孔限制因素,若气孔导度和胞间CO2摩尔分数同时降低,光合作用原料减少,则认为植物净光合速率的下降原因是气孔因素所引起的,相反,叶片净光合速率和气孔导度降低的同时胞间CO2摩尔分数升高,则净光合速率降低的主要原因则为叶肉细胞光合活性的下降所致,即非气孔因素。而本试验结果发现3种玉簪的胞间CO2摩尔分数与净光合速率、气孔导度的日变化呈现相反趋势,在1d中净光合速率、气孔导度的低谷与胞间CO2摩尔分数的高峰同时出现,说明此时限制净光合速率的主导因素正是非气孔限制因素。
光饱和点和光补偿点分别代表植物光合作用之光照强度的上限和下限,同时也代表植物叶片对强光和弱光的利用能力[18]。根据 Boardman[19]的观点,耐荫植物的光补偿点小于20 μmol·m-2·s-1,光饱和点为 50~ 1 000 μmol·m-2·s-1或更低;杜占池和杨宗贵[20]认为光饱和点在500~1 300 μmol·m-2·s-1,光补偿点在 16.7~ 33.3 μmol·m-2·s-1,则为栽培C3植物特征。试验研究得到3种玉簪光饱和点在776.24~802.580 μmol·m-2·s-1、光补偿点在 17.50~ 19.68 μmol·m-2·s-1,这些特征则表明3种叶色玉簪属于耐荫的栽培C3植物类型,但不同叶色玉簪品种其耐荫程度有所不同,其中金头饰玉簪表现出具有较强耐荫能力的C3型植物特征,同时其水分利用效率也相对较高,表明其抗旱能力也较强;黄金叶玉簪与金鹰玉簪则次之,可见,繁殖与应用玉簪属植物应根据品种选择适宜的环境,以利于该植物生长和存活。
从光适应性和水分利用效率两方面进行比较,笔者认为金头饰玉簪对环境的适应性明显优于其他两种玉簪,推广应用前景好,而黄金叶玉簪与金鹰玉簪对环境要求相对苛刻些。
[1]贾洪革.玉簪品种的引种、栽培和组织培养[M].北京:中国科学院植物研究所,2000.
[2]刘接卿,王翠芳,邱明华,等.玉簪花的抗肿瘤活性甾体皂苷成分研究[J].中草药,2010,41(4):520-526.
[3]Meyer M.In vitro propagation of Hosta sieboldiana[J].Hort-Science,1980,15(6):737-738.
[4]施爱萍,张金政,张启翔,等.不同遮荫水平下4个玉簪品种的生长性状分析[J].植物研究,2004,24(4):486-490.
[5]叶子飘.光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J].生态学杂志,2007,26(8):1323-1326.
[6]孙磊,章铁,李宏开,等.柿树光合速率日变化及其影响因子的研究[J].北方果树,2006(2):34-39.
[7]金鑫,胡万良,丁磊,等.遮阴对红松幼苗生长及光合特性的影响[J].东北林业大学学报,2009,37(9):12-13,47.
[8]陶俊,陈鹏,佘旭东.银杏光合特性的研究[J].园艺学报,1999,26(3):157-160.
[9]李映雪,谢晓金,李永秀,等.3种彩叶植物光合生理特性的差异[J].东北林业大学学报,2009,37(5):64-66.
[10]韩清芳,贾志宽,王俊鹏,等.黄土高原地区紫花苜蓿不同叶位光合日变化特征研究[J].草地学报,2009,17(5):558-563.
[11]柯世省,金则新,陈贤田.浙江天台山七子花等6种阔叶树光合生态特性[J].植物生态学报,2002,26(3):363-371.
[12]林夏珍.遮光对窄头橐吾形态及光合特性的影响[J].浙江林学院学报,2008,25(5):614-618.
[13]丁小球,胡玉佳,王榕楷.三种草坪草净光合速率和蒸腾速率的日变化特点研究[J].草业科学,2001,18(2):62-66.
[14]陶汉之.茶树光合日变化的研究[J].作物学报,1991,17(6):444-452.
[15]许大全,丁焕根,苏丽英,等.红豆草和苜蓿的光合效率比较研究[J].生态学报,1991,11(1):89-91.
[16]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[17]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317 -345.
[18]王刚,孙广玉.不同紫花苜蓿品种光合能力的比较[J].东北林业大学学报,2007,35(4):19-21.
[19]Boardman N K.Comparative photosynthesis of sun and shade plants[J].Annual Review of Plant Physiology,1977,28:355 -377.
[20]杜占池,杨宗贵.草原植物光合生理生态研究[J].中国草地,1999(3):20-27.
