基于光照强度变化的毛竹扩张对杉木影响的探讨
2011-05-30周国模白尚斌
刘 烁,周国模,白尚斌
(浙江农林大学 环境与资源学院,浙江 临安 311300)
毛竹Phyllostachys pubescens属于禾本科Poaceae竹亚科Bambusoideae刚竹属Phyllostachys,是中国重要的经济竹种[1],也是中国森林资源的重要组成部分[2]。毛竹依靠强大的根鞭繁殖能力,其面积会随时间的推移不断扩大,且多形成纯林[3],因此,往往会成为一种 “局部入侵”物种[4]。物种入侵会对当地尤其是自然保护区内的生物多样性产生极为不利的影响[5],其影响机制也是科研工作者关注的重点。目前,对于入侵物种众多的研究表明:入侵物种将会从多方面对其他物种产生影响,涉及化感作用[6-8]、竞争[9]、改变环境[10]等。光作为地球上的一切生命过程最基本的能量来源[11],在森林群落形成过程中光环境必然会发生相应变化,影响其他植物生长发育[12-13]。森林群落优势种或建群种的扩散定居导致相邻群落下层光环境变化,继而影响其他植物的生长则是一个值得关注的重要问题。有关植物扩散引起群落下层光环境的变化对森林群落更新影响的研究还鲜见报道。本研究试图从森林群落下层光环境入手,探索毛竹林扩张后光照强度变化对杉木Cunninghamia lanceolata及其幼苗可能存在的影响。
1 研究方法
1.1 研究地概况
本研究以天目山国家级自然保护区已有样带为基础。 保护区位于 30°18′30″~ 30°21′37″N, 119°24′11″~119°27′11″E;年平均气温为8.9~15.8℃,最冷月气温2.6~3.4℃,极端最低气温-20.2℃,最热月气温19.9~28.1℃,无霜期209~235 d;年降水量为1390~1870mm,整个环境条件有利于毛竹的生长。样带为西北—东南走向,设置在毛竹林与针阔混交林过渡地区,宽10 m,长度依照针阔混交林与毛竹纯林交错区域宽度而定。在样带内设定3块10 m×10 m区域作为本次研究的样地。样地各自所包含的林分类型分别为针阔混交林,针阔-毛竹混交林和毛竹纯林,以代表毛竹林蔓延过程中的3种林分状况。经过初步勘察,样地内的针叶树种主要为杉木,3块样地内胸径达到5 cm以上树种以及毛竹数量基本情况如表1。
1.2 光照强度测定
在7月初至9月末的植物生长旺季,利用手持照度计同时测定3块样地内1.5 m高处光照强度,30个点·样地-1,测定时间为8:00-17:00,隔1 h同步测定3组数据,共采集90个数据。在整个观测时期内,间隔7 d测定1次数据。同时,测定空旷地段的光照强度。
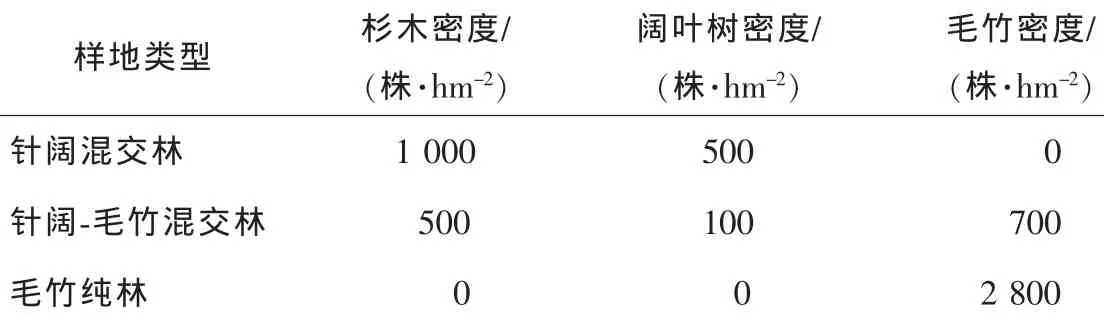
表1 样地内树种组成Table1 Species composition within plots
1.3 毛竹、杉木及其幼苗光补偿点测定
利用Li-6400便携式光合仪于1 d中光合作用较强的时间(9:00-10:00)测定植物的光响应曲线,以计算毛竹、杉木及其幼苗的光补偿点。具体测定方法如下:选择样地内毛竹(2度竹,平均高度11.2 m),杉木(平均胸径15.3 cm,平均树高9.5 m)及其幼苗(平均高53.0 cm,地径1.5 cm)的待测植株各3株,选取位于植株中部向阳面完全展开的叶片5~10片·株-1,选取较为完整的叶片进行测定,测定3~5次·株-1,最后选取相同光照条件下多次测定的平均值,作该植物种类的光合速率曲线。
从光合曲线可以得到植物的最大净光合速率,进而可以计算光补偿点(LCP)。计算公式如下:光补偿点 =Pmaxln(C0)/α。Pmax代表最大净光合速率,α为弱光下光量子利用效率(即表观量子效率),C0为一度量弱光下净光合速率趋近于0的指标[14]。
1.4 数据处理
数据处理利用SPSS以及Excel软件进行。
2 结果与分析
2.1 3种林型内光照强度日变化
3种林分内光照强度以7月4日为例,其单日变化如图1所示。3种林分中,针阔混交林内的光照强度最弱,针阔-毛竹混交林内较毛竹纯林内稍弱,毛竹纯林内光照强度则在3种林分类型内最高。在针阔混交林内光照强度1 d的平均值为13.66μmol·m-2·s-1,对比空旷处降低了85.70%,针阔-毛竹混交林内光照强度(42.24μmol·m-2·s-1)比空旷处降低了 55.91%, 而毛竹纯林内的光照强度平均值为 48.57μmol·m-2·s-1,比空旷处降低49.31%。针阔混交林内光照强度低于针阔-毛竹混交林内的28.59μmol·m-2·s-1,也低于毛竹纯林内的 34.91μmol·m-2·s-1。17:00,最后 1次监测时,针阔混交林内的光照强度为 2.43μmol·m-2·s-1左右。 毛竹纯林内的光照强度仅在 10:00 的监测中(36.70μmol·m-2·s-1) 略低于同时期针阔-毛竹混交林内的光照强度(39.70μmol·m-2·s-1)。在之后的监测中,毛竹纯林内的光照强度则一直高于其他2种林分类型。
由图1可知:光照强度在3种林分类型内的单日变化情况接近于双峰曲线,3种林分内的光照强度从8:00开始缓慢上升,11:00左右达到第1个峰值,之后逐渐下降,在15:00时再次上升,于16:00达到第2个峰值,这在针阔混交林的日变化曲线上表现较为明显。这一情况与空旷地带的单峰变化情况是不同的。造成该现象的原因可能是,在第1个峰值处,阳光处于直射位置,林内光照强度达到最高。这与在空旷地带出现的情况相似,随着太阳高度的变化,阳光的入射角度发生相应改变,到达16:00时,阳光避开了3种林内上层树冠的遮蔽,从林内间隙直接照射林地下层,以一定角度直接照射在监测点上,因此,出现第2个峰值这一情况在空旷地带则没有出现。
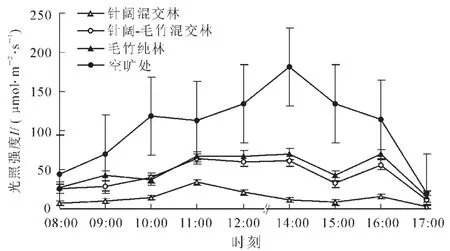
图1 7月4日3种样地类型内光照强度变化及标准误差Figure1 Light intensity variation and standard error in different plots in July 4
2.2 3种林分类型连续多日光照强度变化
如图2所示,在整个监测时期内,毛竹纯林内日平均光照强度始终最高,其日平均最高值(7月 20 日)达到105.80μmol·m-2·s-1,其次是针阔-毛竹混交林(最高值为105.38μmol·m-2·s-1), 最低为针阔混交林(36.77μmol·m-2·s-1), 但其他时期未出现如此高值的原因可能与山区内多为多云天气有关。对于针阔混交林,除7月20日有较为明显的光照强度跃升外,其他监测时期内基本处于平稳态势,变化幅度并不非常明显。但其他2种类型林分内部的光照强度则始终处于波动变化过程中,因为毛竹相较于硬质的针阔树种更易受风的影响产生摆动,使光在某一短时期内可以直接照射到林内,若发生在监测时刻,则使监测数据升高。
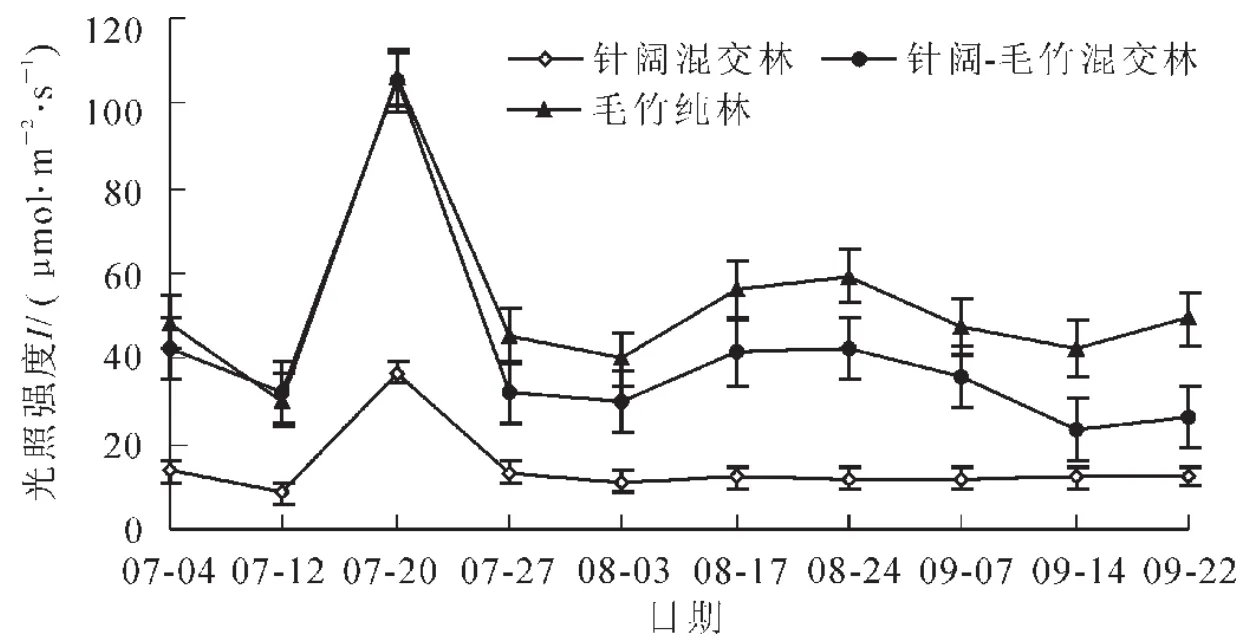
图2 不同样地内光照强度连续日变化曲线及标准误差Figure2 Curve of light intensity and standard error in consecutive days in different plots
通过进一步回归分析,针阔混交林、针阔-毛竹混交林以及毛竹纯林3种林分内光照强度之间存在有如下相互关系:I针阔=0.0273 I针阔-毛竹+0.0320 I毛竹+2.2970。其中I代表光照强度,经过检验,相关系数R达到0.959(R2=0.920),具有较好的相关性。
2.3 毛竹、杉木及其幼苗光补偿点
利用软件Photosyn Assistant绘制毛竹、杉木及其幼苗的净光合速率(图3)。毛竹及杉木的光合速率曲线较为相似,杉木幼苗则与成年树略有不同,随着光照强度的改变成年树种较其幼苗对该变化更加敏感。毛竹在光照强度为0~400.00μmol·m-2·s-1时净光合速率增加较快,之后趋于平稳,而杉木则在400.00μmol·m-2·s-1处趋于平稳,杉木幼苗则几乎在整个测定光强内都在缓慢上升,仅在600.00μmol·m-2·s-1处上升趋势变得缓和。
图3 样本树种光合速率曲线Figure3 Photosynthetic rate curves of the sample trees
由光合速率曲线,可以分别计算出毛竹、杉木、杉木幼苗的光补偿点分别为19.60,9.77,9.78μmol·m-2·s-1。可见,毛竹的光补偿点值远远高于其他的树种。杉木及其幼苗具有相似的光补偿点。结合之前监测所得到的3种林分类型内的光照强度可知,在针阔混交林内出现的日平均最低值为9.85μmol·m-2·s-1,在这一光照强度下,杉木及其幼苗已经能够充分的进行光合作用,并达到光补偿点之上。亦即,在针阔混交林内,杉木及其幼苗可以有效的积累有机物质。再根据图2可得出,针阔-毛竹混交林以及毛竹纯林内的光照强度高于针阔混交林,因此,2种林分类型内的光照强度都能满足杉木的生长,但是杉木幼苗时期喜荫[15],而这2种林分类型内由于遮蔽不及针阔混交林,造成林内直射光照强度较高,因此可能不适合杉木幼苗的生长。
3 结论与讨论
本研究通过对针阔混交林、针阔-毛竹混交林以及毛竹纯林3种林分类型内光照强度的监测,同时测定杉木及其幼苗光补偿点,得到如下的结论:3种林分类型内的光照强度为毛竹纯林>针阔 +毛竹混交林>针阔混交林;通过长时间的监测,杉木及其幼苗的光补偿点皆低于3种林分内长期的光照强度均值,理论上,杉木及其幼苗都可以在3种光照强度下顺利进行光合作用,制造和储存有机物以抵消呼吸作用所带来的消耗。但是,杉木幼苗由于喜荫要求,在毛竹林内较强的直射光照强度下可能会受到一定的抑制,不利于正常生长。
毛竹是重要的经济和碳汇作物,具有的巨大经济价值和生态价值。目前,有关的研究主要还集中在毛竹的丰产栽培技术以及丰产栽培模式等方面,对于它们对生物多样性的不利影响的研究还较少见于报道。因此,适当转变研究视角,将毛竹作为 “局部入侵”物种[4],并对它们对生物多样性产生的影响加以研究,对于保护中国南方森林健康,维持生物多样性具有重要意义。
另一方面,光照强度仅仅是光环境的一个方面。通过监测得到的数据也表明,毛竹入侵带来光照强度的增强并不会导致杉木成年树因光合作用长期低于光补偿点而死亡,因此,这需要在后续的研究中继续探索毛竹林扩张导致的光环境其他方面的变化所带来的影响,同时综合考虑地上部分与地下部分,找到毛竹扩张导致杉木死亡的原因。
[1]宋艳冬,金爱武,金晓春,等.施肥对毛竹叶片光合生理的影响[J].浙江林学院学报,2010,27(3):334-339.SONG Yandong, JIN Aiwu, JIN Xiaochun, et al.Physiology of leaf photosynthesis with fertilization in Phyllostachys pubescens [J].J Zhejiang For Coll, 2010, 27 (3):334-339.
[2]施建敏,郭起荣,杨光耀.毛竹光合动态研究[J].林业科学研究,2005,18(5):551-555.SHI Jianmin, GUO Qirong, YANG Guangyao.Study on the photosynthetic dynamic variation of Phyllostachys edulis[J].For Res, 2005, 18 (5):551-555.
[3]赵明水,刘亮,陆森宏,等.天目山自然保护区毛竹林扩张对植物多样性影响研究[C]//中国林学会.第2届中国林业学术大会:S8野生动物、湿地与自然保护区论文集.北京:中国林学会,2009:190-196.
[4]李伟成,盛海燕,钟哲科.竹林生态系统及其长期定位观测研究的重要性[J].林业科学,2006,42(8):95-101.LI Weicheng, SHENG Haiyan, ZHONG Zheke.Importance of long-term location investigation for bamboo ecosystem[J].Sci Silv Sin, 2006, 42 (8): 95-101.
[5]丁丽霞,王祖良,周国模,等.天目山国家级自然保护区毛竹林扩张遥感监测[J].浙江林学院学报,2006,23(3):297-300.DING Lixia, WANG Zuliang, ZHOU Guomo, et al.Monitoring Phyllostachys pubescens stands expansion in National Nature Reserve of Mount Tianmu by remote sensing [J].J Zhejiang For Coll, 2006, 23 (3): 297-300.
[6]OELRICHS P B, CALANSAN C A, MALEOD J K, et al.Isolation of a compound from Eupatorium adenophorum(Spreng.)[Ageratina adenophora (Spreng.)]causing hepatotoxicity in mice [J].Nat Toxins, 1995, 3 (5):350-354.
[7]宋启示,付昀.紫茎泽兰的化学互感潜力[J].植物生态学报,2000,24(3):362-365.SONG Qishi, FU Yun.Allelopathic potential of Eupatorium adenophorum [J].Acta Phytoecol Sin, 2000, 24 (3):362-365.
[8]TRIPATHI R S, SINGH R S, RAI J P N.Allelopathic potential of Eupatorinum adenophorum: a dominant ruderal weed of Meghalaya India [J].Proc Nat Sci Acad Part B Biol Sci, 1981, 47 (3):458-456.
[9]楼一平,吴良如,邵大方,等.毛竹纯林长期经营对林地土壤肥力的影响[J].林业科学研究,1997,10(2):125-129.LOU Yiping, WU Liangru, SHAO Dafang, et al.Effect of long term management of pure Phyllostachys pubescens stands on soil fertility [J].For Res, 1997, 10 (2): 125-129.
[10]刘红卫,林志凌,苏华轲,等.广东省外来物种入侵现状及其生态环境影响调查[J].生态环境,2004,13(2):194-196.LIU Hongwei, LIN Zhiling, SU Huake, et al.Investigation of the present intrusion situation, and the influence of intrusive alien species on ecology in Guangdong [J].Ecol Environ Sci, 2004, 13(4): 194-196.
[11]牛翠娟,李庆芬,孙儒泳.基础生态学[M].北京:高等教育出版社,2002.
[12]沈荔花,李国庆,戎红,等.入侵植物加拿大一枝黄花对小麦光合生理的影响[J].中国农学通报,2009,25(14):252-255.SHEN Lihua, LI Guoqing, RONG Hong, et al.Effect of invasive plant Solidago canadensis L.on photosynthetic physiology of wheat[J].Chin Agric Sci Bull, 2009, 25 (14): 252-255.
[13]胡天印,方芳,郭水良,等.外来入侵种加拿大一枝黄花及其伴生植物光合特性研究[J].浙江大学学报:农业与生命科学版,2007,33(4):379-386.HU Tianyin, FANG Fang, GUO Shuiliang, et al.Comparison of basic photosynthetic characteristics between exotic invasive weed Solidago canadensis and its companion species [J].J Zhejiang Univ Agric Life&Sci, 2007, 33 (4):379-386.
[14]张小全,徐德应.杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化[J].林业科学,2000,36(3):19-26.ZHANG Xiaoquan,XU Deying.Seasonal changes and daily courses of photosynthetic characteristics of 18-year-old Chinese fir shoots in relation to shoot ages and positions within tree crown [J].Sci Silv Sin, 2000, 36 (3):19-26.
[15]龚垒.树木的光合作用与物质生产[M].北京:北京科学技术出版社,1989.
