C57BL/6J小鼠脂肪组织总RNA提取及周脂素基因的克隆
2011-02-03卫兵艳刘田福郭永昌张引红
卫兵艳,刘田福,郭永昌,张引红
(山西医科大学实验动物中心,太原 030001)
脂肪组织不仅是机体的能量储存库,还是体内很大的内分泌器官;它可以合成或分泌多种具有广泛生物活性的蛋白质或细胞因子,包括周脂素(perilipin)、脂联素(adiponectin)、瘦素(leptin)、肿瘤坏死因子α(TNF-α)、白介素6(IL-6)[1-2]等这些因子与许多肥胖相关性疾病如胰岛素抵抗、糖尿病、内皮功能障碍、高血压病、动脉硬化症等有关。体内能量代谢的核心反应是脂肪细胞的脂解。近年的研究表明[3],该反应主要由脂酶从胞液易位至细胞内脂滴的表面来调控的,该脂滴表面包被的蛋白中周脂素的磷酸化是HSL易位所必需的。由于周脂素在脂质分解中的重要作用,故成为近年来研究的热点。
周脂素是一种新近在大鼠附睾脂肪组织中发现并命名的高磷酸化蛋白,特异表达于脂肪细胞和类固醇生成细胞脂质小滴表面[4]。新近研究表明,脂肪分解调控中的关键蛋白perilipin可能和2型糖尿病、肥胖、动脉粥样硬化等多种代谢性疾病及心血管疾病有关[4-6]。但其作用机制尚不完全清楚。虽然周脂素的功能研究取得了一定进展,但关于周脂素的作用机制以及重组基因药物产品开发等方面,还有待进一步研究。
以脂肪组织为材料提取高质量的总RNA主要存在两方面的问题:第一,脂肪组织RNA含量较少,每毫克组织样品所得总RNA的量小于0.05μg;第二,内源和外源性RNA酶对RNA的降解。因而所提RNA的得率较低。为获得高质量的RNA制品,本实验以脂肪组织为材料,经反复试验,在商家提供的总RNA提取试剂的操作步骤基础上优化和改进了某些提取过程,从而得到了符合实验要求的高质量的总RNA。利用改进后的方法提取了C57BL/ 6J小鼠脂肪组织RNA。以此为模板进行RT-PCR,克隆了周脂素基因并对核酸序列进行分析,为下一步进行重组周脂素的大量表达及其生物学功能研究奠定基础。
1 材料和方法
1.1 材料
1.1.1 实验动物:C57BL/6J小鼠购于山西医科大学实验动物中心,生产许可证号:SCXK(晋2009-0001)。实验过程中对动物处置符合2006年科学技术部发布的《关于善待实验动物的指导性意见》[7]。
1.1.2 载体及宿主菌:受体菌E.coli JM109由本室保存;pMD18-T购自大连TaKaRa公司。
1.1.3 其他相关试剂:RNAisoTMPlus、限制性内切酶SpeⅠ和NotⅠ、DNA marker,均购于大连TaKaRa公司;质粒抽提试剂盒购于 O MEGA公司,LAmp二步法 R T-PCR试剂盒、DNA回收纯化试剂盒、T4DNA连接酶、加A试剂盒均购于北京康为世纪生物科技有限公司。
1.2 方法
1.2.1 总RNA的提取:按照试剂盒说明书进行,部分操作步骤做相应的改进:(1)取新鲜的脂肪组织并将样品的量加大到1 g,RNAisoTMPlus的量加大到20 m L。(2)第1次离心后上层含有大量油脂,用吸头除去,保证完全去除油脂污染,然后再离心5 min。(3)进行两次氯仿抽提,以去除DNA的污染。(4)用75%的乙醇洗RNA沉淀后,再用无水乙醇洗1次,缩短RNA沉淀晾干的时间,可避免RNA在空气中暴露时间过长而增加外源RNase对RNA的降解。紫外分光光度计测定RNA的纯度及含量,利用琼脂糖凝胶电泳快速鉴定所提取的总RNA的质量。
1.2.2 引物设计、反转录-聚合酶链反应及其产物纯化
1.2.2.1 引物设计与合成:根据GenBank中C57BL/6J小鼠周脂素的基因的序列,设计引物:序列如下(下划线部分为酶切位点,斜体部分为保护性碱基。):5'端引物:5'-GGACTAGTCTTGGGCG TTTGCCTTACCT-3'(引入 S peⅠ酶切位点及保护性碱基),3'端引物:5'-AAGGAAAAAA GCGGCCGCCCTTGACGAGCAGCGACCTT-3'(引入NotⅠ酶切位点及保护性碱基)用于扩增周脂素全长编码区。该引物由北京奥科公司合成。
1.2.2.2 RNA逆转录为cDNA:在0.2 m L的PCR管中加入下列成分:总 R NA(0.5μg/μL)4μL,Oligo d(T)18(100μmol/L)1μL,dNTP(2.5 mmol/L each)4μL,灭 R NA酶水4μL,混匀后置PCR仪中,65℃,5 min,迅速冰浴10 m in,短暂离心后继续加入下列成分:5×RT buffer,5μL,RNA酶抑制剂(20 U/μL)1μL,0.1 mol/L DTT 2μL,混匀后置PCR仪中,37℃,孵育2 min后加入HiFi-MMLV逆转录酶 ( 200 U/μL)1μL,混匀后置PCR仪中,37℃50 min,终止反应,置 - 20℃冰箱保存备用。
1.2.2.3 周脂素基因的扩增:在0.2 mL的PCR管中加入下列成分:cDNA 4μL,10×LAmp PCR buffer 5μL,10μmol/L的正、反义引物各 2 μL,dNTP Mix(2.5 mmol/L each)4μL,5×RT-PCR enhancer 10μL,LAmp DNA polymerase 0.5μL加双蒸水补足体积至50μL,混匀后置PCR仪中。PCR反应条件为:94℃预变性2 min,然后94℃变性30 s,60℃退火30 s,72℃延伸60 s,循环30次,72℃延伸5 m in,终止反应。取PCR扩增产物用1.2%琼脂糖凝胶进行电泳,紫外灯下观察电泳结果并照相。
1.2.2.4 PCR产物的纯化回收:PCR扩增产物经1.2%琼脂糖凝胶电泳分离,在紫外灯下切取含有目的基因的琼脂糖凝胶块,按胶回收试剂盒说明书中的方法,回收胶块中的目的片段。
1.2.3 周脂素基因编码区克隆、酶切及其DNA序列分析:
1.2.3.1 PCR产物加A:纯化的目的基因7μL,dATP 1μL,10×A-tailing buffer 1μL,Tag DNA polymerase 1μL,混匀后置PCR仪中,72℃保温30 m in。
1.2.3.2 PCR产物T-A克隆及鉴定:在10μL连接反应体系中,将加A的目的片断与pMD18-T载体按摩尔数3∶1混合,加入T4DNA连接酶和连接缓冲液,22℃孵育1 h后,65℃10 m in灭活T4DNA连接酶,然后取上述连接产物10μL转化大肠杆菌DH5α感受态细菌100μL,将转化后的DH5α涂布于含氨苄青霉素(100μg/m L)及X-Gal(40 μg/m L)、IPTG(24μg/m L)的LB固体培养基平板上,37℃培养过夜,挑取单个白色菌落置于LB液体培养基中,扩大培养后提取质粒,经speⅠ/NotⅠ双酶切鉴定含有目的基因的质粒由大连TaKaRa公司进行DNA测序,并将测序结果与GenBank中的已知基因序列及氨基酸序列进行比对。
1.2.3.3 C57BL/6J重组质粒的DNA序列分析:阳性克隆用通用引物测序,测序结果表明序列与GenBank比对达到100%一致(GenBank accession number:NM_175640.2)。
2 结果
2.1 C57BL/6J小鼠附睾脂肪组织总RNA的提取结果
提取的总RNA经紫外分光光度仪测定A值。A260/A280=1.89,比值在1.8~2.0之间;总RNA经1%琼脂糖凝胶电泳可见28、18s 2条清晰的条带,并且28s/18s≈2.0(图1),表明提取的总RNA质量符合后续实验的要求,可用于逆转录。

图1 C57BL/6J脂肪组织总RNA琼脂糖凝胶电泳Fig.1 Representative electrophoresis tracing of the total RNA from C57BL/6Jmouse adipose tissue
2.2 RT-PCR结果
提取C57BL/6J小鼠附睾脂肪组织的总RNA,经RT-PCR扩增后,1.2%琼脂糖凝胶电泳可见与预期设计的周脂素基因片段大小一致的扩增条带(包括两侧酶切位点和保护碱基)(图2)。

图2 反转录-聚合酶链反应产物琼脂糖凝胶电泳Fig.2 Representative electrophoresis tracing of the RT-PCR products
2.3 PCR产物T-A克隆结果
经RT-PCR扩增的周脂素基因片段与pMD18-T载体连接构建成pMD18-T-plin重组载体,经speⅡNotⅠ双酶切,1.2%琼脂糖凝胶电泳可见2条带,中1条带与RT-PCR产物的电泳结果完全一致,另1条与pMD18-T载体片段(2 692 bp)的大小一致,见图3。
2.4 周脂素重组质粒的DNA序列分析
阳性克隆用通用引物测序,测序结果表明序列与GenBank比对均为100%一致(GenBank accession number:D45371)。测序图谱见图4、5。表明pMD18-T载体中已含有周脂素基因片段。

图3 周脂素重组质粒酶切鉴定结果Fig.3 Electrophoresis of the recombinant plasm id perilipin using double enzyme digestion
3 讨论
由于脂肪组织富含脂类物质,单位体积细胞数较少,且单位细胞RNA含量也相对较少,按照RNAisoTMPlus试剂说明书提取脂肪组织RNA效果并不理想,所以如何有效地除去脂类的污染和最大限度地提高产量是在提取脂肪组织RNA过程中不可忽视的问题。在脂肪组织RNA提取过程中,对其中一些步骤作了适当的改进。本实验主要做到以下几点:①样品量足,一般组织的RNA提取样品量大概是100 mg,本实验将样品量加大到1 g进行提取从而保证了RNA的得率。②样品组织裂解充分,短时间内液氮研磨至粉末状加入超量的裂解液快速研磨至透明后剧烈震荡3 min,保证了裂解液与组织的充分接触。③除脂彻底,将样品与裂解液反应后,离心5 min后,用吸头除去了上层的脂类,取得的下层溶液应做同样的2次离心以保证完全去除脂类污染。④去除内源和外源性RNA酶的影响,实验过程中使用的实验材料均用0.1%的DEPC水处理后,高压灭菌,操作过程戴口罩帽子,尽量保持环境洁净,液氮研磨充分,用75%的乙醇洗完RNA沉淀后,再用无水乙醇洗1次[8],以此减少内外源性RNA酶的影响。从而得到了高质量的RNA,为后续扩增周脂素基因做了充分保证。
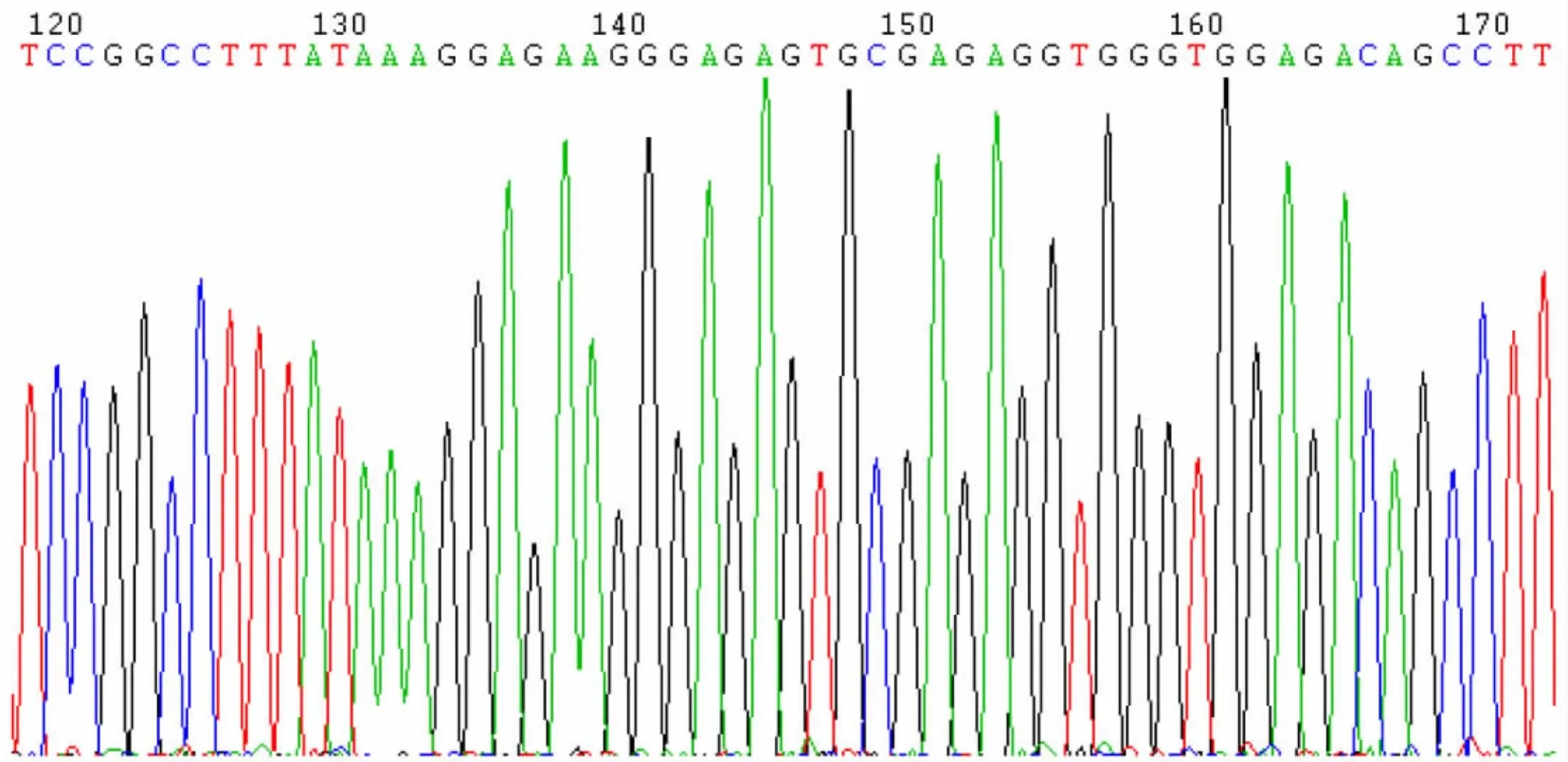
图4 周脂素的测序图谱Fig.4 Sequencing map of the plasm id perilipin

图5 周脂素克隆序列比对部分结果Fig.5 Sequence analysis of the plasm id perilipin
周脂素由Plin基因编码[9]。Perilipin氨基末端大约100个氨基酸的区域与ADRP的氨基末端区域极为相似,被称为PAT结构域。鼠的perilipin基因定位于第7号染色体,人的基因定位于15q26,含有9个外显子。研究发现,perilipin可能在脂肪分解调控中起到“分子开关”的作用,在脂肪水解和机体能量代谢过程中发挥着双重作用[3],既可通过阻止脂肪酶接近脂滴降低基础状态下的脂解,又可促进激素刺激的脂肪分解,可以调控脂解率以及脂肪组织中甘油三酯的输出。肿瘤坏死因子α(TNF-α)、过氧化物酶体增殖物激活受体γ(PPARγ)激动剂、瘦素(leptin)等均可以影响perilipin的表达[10-12]。由于周脂素与脂代谢的具体作用目前还不明确[13,14],其可能是联系胰岛素抵抗与代谢性疾病的重要环节,在脂质沉积及有序排列中发挥重要的调控作用[15-16],其在脂代谢中的关键作用可能与肥胖及相关疾病的发生有关[17]。
实验经过提取脂肪总RNA、Rt-PCR、T-A克隆,得到周脂素编码区重组质粒,测序结果表明,序列与Genbank比对均为100%一致(GenBank accession number:NM_175640.2)。本实验得到没有任何突变的基因,考虑与以下因素有关:①附睾脂肪组织总RNA的提取质量较高为进行反转录提供保证。②引物的GC比例设定合适(上游为55%,下游为60%),因而提高了基因扩增的特异性。③割胶纯化时,操作速度很快,减少了紫外线对目的基因的损伤。紫外线可引起DNA损伤,应尽量减少其照射时间。④对纯化的目的片段进行了加A反应,同时pMD18-T克隆载体是线性化的载体,其3’端有1个T伸出,显著提高了重组效率。本实验成功地克隆没有任何突变的周脂素基因,为对其进行精确实验提供了根本保障;为进一步真核表达周脂素蛋白,建立周脂素转基因小鼠,分析周脂素的生物功能及作用机制提供了实验基础;为进一步理解体脂分布异常,胰岛素抵抗开辟一新的方向,最终为探索该类药物的开发提供崭新的思路;可能也将为糖尿病等相关疾病的临床工作提供一种新的防治策略。
[1]Crossno JT Jr,Majka SM,Grazia T,et al.Rosiglitazone promotes development of a novel adipocyte population from bone marrow-derived circulating progenitor cells[J].J Clin Inves,2006,16(12):3220-3228.
[2]Dalen KT,Dahl T,Holter E,et al.LSDP5 is a PAT p rotein specifically expressed in fatty acid oxidizing tissues[J].Biochim Biophys Acta,2007,1771(2):2102227.
[3]Moore HP,Silver RB,Mottillo EP,et al.Perilipin targets a novel pool of lipid droplets for lipolytic attack by hormonesensitive lipase[J].J Biol Chem,2005,280(52):43109 -43120.
[4]Greenberg AS,Egan JJ,Wek SA,et al.Perilipin,a major hormonally regulated adipocyte2 specific phosphoprotein associated with the periphery of lipid storage drop lets[J].J Biol Chem,1991,266(17):11341-11346.
[5]Souza SC,Vargas LM,Yamamoto MT,et a1.Overexpression of perilipin A and B blocks the ability of tumor necrosis factor alpha to increase lipolysis in 3T3-L1adipocytes[J].JBiol Chem,1998,273:24665-24669.
[6]Londos C,Sztalryd C,Tansey JT,et al.Role of PAT proteins in lipid metabolism.Biochimie,2005,87:45-49.
[7]中华人民共和国科学技术部.关于善待实验动物的指导性意见[S].2006-09-30.
[8]曹廷兵,叶治家,巩燕,等.一种快速鉴定RNA质量的方法[J].第三军医大学学报,2002,24(8):992-993.
[9]Martinez-Botas J,Anderson JB,TessierD,et al.Absence of perilipin Results in leanness and reverses obesity in Lepr(db/ db)mice[J].Nat Genet,2000,26:474-479.
[10]Fauny JD,Silber J,Zider A.Drosophila lipid storage droplet 2 gene(Lsd-2)is expressed and controls lipid storage in wing imaginal discs[J].Dev Dyn,2005,232(3):725-732.
[11]Londos C,Brasaem le DL,Schultz CJ,et al.On the control of lipolysis in adipocytes[J].Ann N Y Acad Sci,1999,892:155 -168.
[12]Arimura N,Horiba T,Imagana M,et al.The peroxisome proliferator 2 activated receptor gamma regulates expression of the perilipin gene in adipocytes[J].JBiol Chem,2004,279(11):10070-10076.
[13]Sztalryd C,Xu G,Dorward H,et al.Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation[J].JCell Biol,2003,161:1093-1103.
[14]Clifford GM,Londos C,Fredric B.et al.Translocation of hormone-sensitive lipase and perilipin upon lipolytic stimulation of rat adipocytes[J].JBiol Chem,2000,275:5011-5015.
[15]Tansey JT,Sztalryd C,Gruia-Gray J,et al.Perilipin ablation Resultsin a lean mouse with aberrant adipocyte lipolysis,enhanced leptin production,and resistance to diet-induced obesity.Proc Natl Acad Sci U S A,2001,98(11):6494 -6499.
[16]Kawai T,Ng MC,Hayes MG,et al.Variation in the perilipin gene(PLIN)affects glucose and lipid metabolism in non-Hispanic white women with and without polycystic ovary syndrome.[J].Diabetes Res Clin Pract.2009,86(3):186 -92.
[17]Kern PA,Di Gregorio G,Lu T,et al.Perilipin expression in human adipose tissue is elevated with obesity[J].J Clin Endocrinol Metab,2004,89(3):1352-1358.
