脊尾白虾的行为学观察研究
2010-10-23万夕和姚国兴陈爱华张志勇王李宝吴国军张曹进刘海林
沈 辉, 万夕和, 许 璞, 姚国兴, 陈爱华, 张志勇, 王李宝, 吴国军,张曹进, 刘海林
(1. 江苏省海洋水产研究所, 江苏 南通 226007; 2. 常熟理工学院, 江苏 常熟 215500)
脊尾白虾的行为学观察研究
沈 辉1, 万夕和1, 许 璞2, 姚国兴1, 陈爱华1, 张志勇1, 王李宝1, 吴国军1,张曹进1, 刘海林1
(1. 江苏省海洋水产研究所, 江苏 南通 226007; 2. 常熟理工学院, 江苏 常熟 215500)
在高密度室内饲养条件下, 观察了脊尾白虾(Exopalaemon carinicauda)的摄食、相残、运动、蜕皮、间隔、攻击和防御行为。试验结果表明, 脊尾白虾摄食行为活跃, 运动行为多样, 在饥饿或个体较大差异条件下具有一定程度的相残行为, 蜕皮行为表现出一定的特殊性, 常经数天多次蜕皮行为特征后才能将旧壳蜕下, 间隔行为不明显, 攻击频率不高。另外, 通过设置不同投喂组和投喂间隔时间观察比较了索饵率和相残率, 结果表明, 索饵率在第 8小时已基本达到最高, 相残率随投喂间隔时间的延长和空间密度的增大而升高。
脊尾白虾(Exopalaemon carinicauda); 摄食行为; 相残行为; 蜕皮行为; 防御行为
脊尾白虾(Exopalaemon carinicauda), 又名小白虾, 系热温带海区底栖虾类, 具有繁殖能力强(1 a内可以多次产卵且可以在养殖池中自繁)、生长速度快(1 a内可以多茬养殖收获)、环境适应能力强(广温、广盐, 广食)等优点[1,2]。随着沿海滩涂的开发和大面积水产养殖业的发展, 脊尾白虾的人工养殖规模也在逐年扩大, 且已成为沿海滩涂地区主要的特色水产品。脊尾白虾作为新兴的养殖品种, 其研究报道还较少, 在已报道的研究中主要集中在其生物学和养殖繁育技术[3~8]。在目前的养殖模式下, 脊尾白虾的养殖密度很高, 其养殖环境较之自然生长环境发生了很大变化, 其生长、发育及繁衍等行为会因生存环境的改变而发生变化, 甚至会缺失某些行为, 导致应激反应、生长发育及免疫力等方面的下降。迄今,脊尾白虾行为生态学方面的研究还尚未见报道。因此, 作者于2009年5~6月在江苏省海洋水产研究所繁育基地高密度养殖条件下进行了脊尾白虾的行为习性观察研究。
1 材料和方法
1.1 实验用虾
实验用脊尾白虾取自南通启东吕四养殖场, 取养殖中期虾: 虾体长(3.5±0.4) cm, 1 000尾, 体质量为(2.1±0.3) g, 健康, 无伤病。
1.2 实验方法
在实验水池中暂养5 d后, 以3个空间密度(100、200和400尾/m3)转入观察水槽(1.5 m×0.6 m×0.6 m),并设置平行对照组。水温 24℃±1℃, 盐度 30.2±0.5,充气泵间隔充气并保持DO>5.0 mg/L。通过录像对其摄食、相残、运动、间隔、蜕皮、攻击和防御等行为进行观察研究。以400尾/m3设置组为观察组, 比较投喂对虾配合饲料与沙蚕(Nereis succinea) 在5、30 s、2、10及30 min内上、中、下各水层的分布比例; 另外, 将投喂饲料间隔时间设置为 1、3、8、16及 24 h, 通过记录 10 min内对虾抱食饲料的比例,比较其索饵率。3个密度梯度设置组投喂间隔时间设置为1、3、8、16、24、48 h, 统计各过程中的相残率。作详细记录, 连续观察15 d。
2 结果
2.1 摄食行为
投喂对虾配合饲料后, 脊尾白虾反应积极, 步足游动速度加快, 迅速游到饲料上方, 用颚足将颗粒抱起, 大颚磨碎食物, 小颚咀嚼食物。部分饲料漂浮于水面, 脊尾白虾可以采用仰泳的姿势将饲料抱于怀中进行啃食。部分小个体虾用步足抱持饲料颗粒后并不立即伏于水底啃食, 而是环绕水槽游动几周后等虾群基本都处于啃食饲料状态后才伏于水底或以游动的方式进行啃食饲料颗粒。部分个体较大的虾常用步足抱持两粒饲料, 伏于水底后将其中一粒进行啃食, 将另一粒放于颚足控制范围内。另外,大部分虾会将饲料颗粒全部啃食吃下, 但有些虾将饲料颗粒啃食部分后丢弃, 丢弃的饲料会被另外的虾捡起后继续啃食, 而被啃食后的饲料常会很快再次被丢弃。
通过设置不同的投喂间隔时间比较脊尾白虾的索饵强度, 结果表明, 在前8 h内, 索饵强度随着时间的延长相应地增强(图1)。但8与16、24 h的索饵率已几乎无差别, 表明在脊尾白虾的饲养中8 h为较合理的投喂间隔时间。

图1 脊尾白虾不同投喂时间的索饵率Fig. 1 The rate of feeding of Exopalaemon carinicauda at different times after the proceeding feeding
投喂小尺寸沙蚕时, 脊尾白虾静置沙蚕旁边约2~5 s后突然用其步足将沙蚕抓住, 且用其第一、二对步足将沙蚕身体向前拉直, 控制住沙蚕的身体让其无法扭动, 然后将沙蚕直接送入口中咀嚼。投喂大个体沙蚕时, 脊尾白虾无法抱持住扭动的沙蚕, 几轮尝试捕食后, 脊尾白虾放弃捕食沙蚕的行为。将沙蚕肢解后再次投于水槽中, 脊尾白虾反应积极, 迅速游动抢食沙蚕组织, 小组织块被脊尾白虾抱持后环绕水槽游动, 待虾群平静后独自啃食, 而大组织块会被脊尾白虾包围后共同用大颚撕扯、切割, 进而用小颚咀嚼沙蚕。另外, 本实验将沙蚕组织液滴入水槽观察脊尾白虾的行为反应, 发现组织液滴入水槽后5~10 s后, 虾群表观出强烈的索饵行为。
通过记录不同时间段上、中、下水层虾个体的数量, 比较索饵的强度, 结果发现, 投喂饲料组与投喂沙蚕组在第30秒时, 上层和中层虾的数量比例都是最大, 表明脊尾白虾在第30秒时索饵强度基本达到最大, 继而过渡为伏底摄食行为(图2)。由5、30 s,2、10、20 min 5组不同投喂组各水层所占百分率发现, 投喂沙蚕组的索饵强度要高于投喂饲料组, 且索饵持续时间也要长于后者。另外, 从两组第10与20分钟各水层所占率来看, 脊尾白虾在第10分钟基本处于静伏水底啃食饵料状态, 索饵行为过渡为摄食行为。
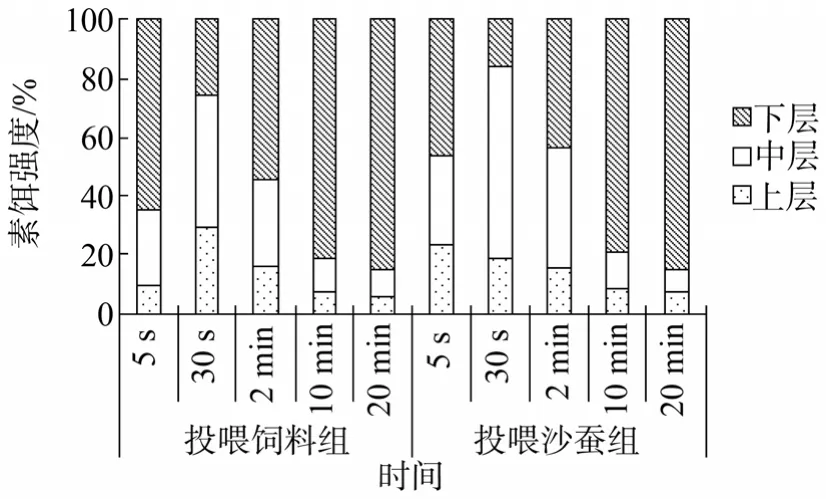
图2 不同投喂组不同时间各水层分布百分率Fig. 2 The percentage of Exopalaemon carinicauda in different water phases and feeding sets
2.2 相残行为
观察发现脊尾白虾具有一定程度的相残行为。脊尾白虾的相残率随着投喂间隔时间的增长及设置密度的增大而增高(图3)。饵料不足或空间密度较大时, 脊尾白虾会捕食刚蜕皮的虾。一般是大个体虾捕食小个体虾, 大个体虾从小个体虾背后突然用步足抱住小个体虾的头胸部, 然后将其步足甚至游泳足全部咬断, 然后从头胸部和腹节的连接处开始啃食,被啃食剩下的个体组织还会被其他虾捡起后继续啃食。也观察到几只小个体虾同时攻击一只大个体虾,最后将大小体虾共同啃食的现象。残食刚蜕皮虾时,通常将虾组织全部啃食, 而未处于蜕皮期的虾个体组织被残食后通常剩下头胸部。

图3 不同投喂组不同投喂时间的相残率Fig. 3 The cannibalism percentage of Exopalaemon carinicauda at different feeding times and sets
2.3 运动行为
脊尾白虾在游动时, 将两个触角分别置于身体两侧, 步足收拢于头胸部, 游泳足不停地划水。脊尾白虾较为安静, 大部分时间伏于槽底, 常在槽底和中下水层做间歇性游动。有时脊尾白虾突然以约 45度角斜向上迅速游动, 到达水面后转以仰泳方式运动。当换水、投喂饵料及搅到池水等情况时, 脊尾白虾虾群会出现集体环水槽游动的行为。
2.4 攻击和防御行为
脊尾白虾的攻击行为主要表现为额剑撞击和步足抓挠两种方式。额剑撞击主要发生在游动过程中大个虾对小个体虾的攻击, 而步足抓挠行为主要发生在摄食过程中两个虾的规格比较一致时, 双方使用步足抱持在一起相互抓挠, 通常抓挠行为持续2~4 s就相互分开。
脊尾白虾的防御行为主要为避让和尾扇向后弹开两种方式。小个体虾在游泳过程中在触角控制的范围内遇到大个体虾会采取主动避让的方式躲避大个体的攻击。尾扇向后弹开使虾体急速向后是脊尾白虾最重要且最为有效的防御行为, 主要发生在小个体虾遭遇攻击时、两虾相互抓挠攻击后弱者溃败时及刚蜕皮虾遭遇其他虾的触碰时。
2.5 蜕皮行为
脊尾白虾的蜕皮行为主要发生在夜晚或早晨,但光线不足的午后也会发生蜕皮行为。蜕皮刚开始时, 脊尾白虾摄食量减少, 且运动行为减弱, 虾体大量吸收水分, 使身体膨胀, 头胸甲出现明显的透明接缝。蜕皮时, 脊尾白虾伏于水槽底部, 虾体开始间歇性地弯曲。随着弯曲次数的增加, 其腹节抖动的程度也随之增强, 最剧烈时虾的头胸部, 触角、腹节及腹肢同步地抖动, 腹节约弯曲 8~10次, 头胸甲与腹节交接处背膜裂开, 且第四、五腹节交界处也出现开裂, 该行为持续约1 h后对虾开始无方向性地剧烈游动, 最后在水槽底部骤然跳跃, 个体从旧壳的背膜处跃出。刚蜕皮虾个体步足收拢不能展开, 且无法支撑身体, 靠游泳足和尾扇进行躲避其他虾的攻击。对单只和群体脊尾白虾观察发现, 脊尾白虾通常多次中断身体弯曲的蜕皮状态, 约2~4 d连续出现以上行为后, 才真正地完成蜕皮。
2.6 间隔行为
脊尾白虾的间隔行为并不明显, 个体较小者或个体较为一致的虾并不表现出明显的间隔行为。但大个体虾在其大触角范围内有其他小个体虾时, 则表现出间隔行为, 其大触角开始摆动, 身体也伴随着微微颤抖。通常情况下, 小个体虾选择躲避离开大个体虾的间隔领域, 偶尔大个体虾会立即攻击小个体虾, 用步足抓挠或额剑碰击驱赶小个体虾离开自己的间隔领域。
3 讨论
甲壳动物的蜕皮行为一直伴随着其幼体的变态发育及成体的生长发育的全过程。据研究报道[9~11],不同对虾的蜕皮行为过程较为类似, 但部分细节并不相同, 如蜕皮过程的连续性、过程的长短、腹节弯曲的次数及背膜开裂的部位等方面。王安利等[9]对中国对虾的蜕皮行为观察发现, 中国对虾(Penaeus chinensis)在蜕皮过程中仅通过 4~6次急剧曲伸, 头胸甲背膜即会裂开, 再经过几次骤然上跳, 新体就跃出旧壳, 即完成蜕皮过程。Longmuir[10]在对香蕉虾(Penaeus merguiensis)蜕皮行为观察时发现, 香蕉虾从开始出现腹节弯曲到蜕皮完成需要弯曲约 15次,蜕皮过程即可完成。在本实验多组观察中发现, 脊尾白虾以腹节弯曲为信号的蜕皮行为通常中断, 且在此之后的2~4 d一直出现以上类似的蜕皮行为特征,且蜕皮行为经常中止, 脊尾白虾开始转为正常的其他行为特征, 甚至立即出现摄食行为, 但此时虾的第四与第五腹节处已经出现开裂的痕迹, 如此反复多次后, 会经常观察到多只虾的第四、第五腹节交界处呈红褐色。另外, 不同的虾其蜕皮过程所需的时间也不相同, 香蕉虾[10]与中国对虾[9]从开始腹节弯曲到新体跃出旧壳分别仅需40 s和5 min, 而脊尾白虾通常需要约1 h完成蜕皮过程。虾类的蜕皮周期和行为除受内源因子 (蜕皮激素、Y 器官及 X 器官等)[11~13]的影响外, 还会受到温度、盐度、pH、钙镁离子等因素的共同影响[14~18]。本实验中脊尾白虾多次中断蜕皮行为的机理尚不清楚, 还需要对其内源因子及外源因子等多种影响因素进行系统研究。
甲壳动物的相残行为一直是影响自然界甲壳动物分布密度的主要因素, 也是制约其育苗养殖的瓶颈之一[19]。水生动物相残行为学的研究主要集中在鱼类[20~23]和蟹类[24~25]方面, 对虾类相残的研究相对较少, 仅见中国对虾、凡纳滨对虾(Litopenaeus vannamei)、日本对虾(Penaeus japonicus)、褐虎虾(Peaeus esculentus)等对虾品种[10~11,26~29]。张沛东等[26,27]对中国明对虾(Fenneropenaeus chinensis)与凡纳滨对虾仔虾的相残行为研究发现, 中国对虾仔虾相残行为高发, 攻击频率高, 而凡纳滨对虾仔虾的相残现象少,攻击频率低。甲壳动物的相残行为除了内源因素影响外, 其他因素如幼体密度、个体大小差异及不同饥饿状态等方面也对其成体及幼体的相残行为具有不同程度的影响[10~11,24~28]。陈学雷等[29]在对日本对虾仔虾的相残行为研究中发现, 相残率受饵料密度、个体大小差异、光线条件的影响, 并提出可以通过充气进而降低放养密度从而有效地降低仔虾的间隔行为及攻击行为, 改善仔虾的相残行为。本实验的观察结果证明了饥饿与饲养密度直接影响对虾的自残行为。以此推测, 目前脊尾白虾的养殖主要以粗饲料为主, 其饵料密度较低, 造成一定程度的饥饿进而引发相残。另外, 其虾苗主要以留塘种虾自繁为来源,种虾繁殖的不同步导致群体中个体的大小差异, 直接导致相残行为。
[1] 李明云. 池养脊尾白虾的繁殖、生长及其最大持续轮捕量的初步探讨[J]. 水产学报, 1994, 18(2): 85-92.
[2] 王兴强, 阎斌伦, 马 , 等. 脊尾白虾生物学及养殖生态学研究进展[J]. 齐鲁渔业, 2005, 22(8): 21-23.
[3] 李明德. 天津脊尾白虾Palaemon(Exopalaemon carincauda)个体生态[J]. 现代渔业信息, 2005, 10: 12-13.
[4] 徐加涛, 徐国成, 于斌, 等. 脊尾白虾繁殖生物学及人工育苗生产技术[J]. 中国水产, 2007, 4: 52-55.
[5] 陈卫平. 不同温度、盐度脊尾白虾 Exopalaemon carincauda (Holthuis)早期胚胎和幼体发育的观察研究[J]. 现代渔业信息, 2005, 20(5): 23-26.
[6] 董存有. 珠江口脊尾白虾的一些生物学观察[J]. 四川动物, 1989, 8(4): 36-38.
[7] 董建波, 程建军, 何健. 脊尾白虾 Exopalaemon carinioauda(Holthuis)和三疣梭子蟹 Portunus trituberculatus(Miers)健康养殖技术探讨[J]. 现代渔业信息, 2008, 23(12): 26-29.
[8] 欧海金, 谢汉阳, 夏武强, 等. 梭子蟹与脊尾白虾、贝类生态混养技术[J]. 齐鲁渔业, 2008, 25(10):16-17.
[9] 王安利, 母学全, 周世梅, 等. 中国对虾行为的观察[J]. 海洋科学, 1993. 4: 16-18.
[10] Longmuir E. Setal development, moult-staging and ecdysis in the banana prawn Penaeus merguiensis[J].Marine Biology, 1983, 77: 183-190.
[11] Mc Conaugha J R. Nutrition and larval growth[J].Larval Growth, 1985, 1-66.
[12] Mc Conaugha J R. Identification of the Y-Organ in the larval stages of the crab, Cancer Anthonyi Rathbun[J].J Morph, 1980, 164: 83-88.
[13] 王芳, 穆迎春, 董双林, 等. 去眼柄对凡纳滨对虾稚虾蜕皮和生长的影响[J]. 中国海洋大学学报. 2004,34(3): 371-376.
[14] Keys S J. Aspects of the biology and ecology of the brown tiger prawn, Penaeus esculentus, relevant to aquaculture [J]. Aquaculture, 2003, 217: 325-334.
[15] Keys S J, Crocos P J. Domestication, growth and reproductive performance of wild, pond and tankerreared brown tiger shrimp Peaeus esculentus[J].Aquaculture, 2006, 257: 232-240.
[16] Etienne B, Patrick K, Charles M. Effect of stocking density on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions [J].Aquaculture, 2003, 219: 241-255.
[17] Nga B T, Lűrling M, Peters E T H M, et al. Chemical and physical effects of crowding on growth and survival of Penaeus monodon Fabricius post-larvae[J].Aquaculture, 2005, 246: 455-465.
[18] Vijayan K K, Diwan A D. Influence of temperature,Salinity, pH and light on molting and growth in the Indian white prawn penaeus indicus(Crustacea: Decapoda:Penaeidae) under laboratory conditions[J]. Asian Fisheries Science, 1995, 8: 63-72.
[19] Dall W, Hill B J, Bothlisberg P C, et al. The biology of the penaeidae[J]. Advance in Marine Biology,297-394.
[20] 华元渝, 陈亚芬, 钱林峰. 暗纹东方 苗种同类相残现象的研究[J]. 水生生物学报, 1998, 22(2): 195-197.
[21] Baras E, Maxi M Y J, Ndao M, et al. Sibling cannibalism in dorada under experimental conditions(Ⅱ). Effect of initial size heterogeneity, diet and light regime on early cannibalism[J]. Journal of Fish Biology, 2000,57: 1 021-1 036.
[22] Qin J, Fast A W. Size and feed dependent cannibalism with juvenile snakedhead Channa striatus[J]. Aquaculture, 1996, 144: 313-320.
[23] 邹桂伟, 罗相忠, 潘光碧. 大口鲇苗种同类相残的研究[J]. 中国水产科学, 2001, 8(2): 55-58.
[24] Lovrich G A, Sainte M B. Cannibalism in the snow crab,Chionoecetes opilio(O. Fabricius)(Brachyura: Majidae), and its potential importance to recruitment[J]. J Exp Mar Biol Ecol, 1997, 211: 225-245.
[25] 陈学雷, 李少菁, 王桂忠, 等. 拟穴青蟹大眼幼体相残与资源可获得性和饥饿关系的研究[J]. 厦门大学学报, 2008, 47(1): 99-103.
[26] 张沛东, 张秀梅, 李健, 等. 中国明对虾、凡纳滨对虾仔虾的行为观察[J]. 水产学报, 2008, 32(2):223-228.
[27] 张沛东, 张秀梅, 李健. 对虾的行为生态学研究进展[J]. 应用生态学报, 2006, 17(1): 127-130.
[28] Marcelo A, Eduardo P P, Eucario G L. White shrimp penaeus vannamei culture in freshwater at three densities: condition state based on length and weight[J].Aquaculture, 2008, 283: 13-18.
[29] 陈学雷, 林琼武, 李少菁, 等. 日本对虾仔虾相残的实验研究[J]. 厦门大学学报(自然科学版), 2003,42(3): 358-362.
Behavior of Exopalaemon carinicauda
SHEN Hui1, WAN Xi-he1, XU Pu2, YAO Guo-xing1, CHEN Ai-hua1, ZHANG Zhi-yong1,WANG Li-bao1, WU Guo-jun1, ZHANG Cao-jin1, LIU Hai-lin1
(1. Institute of Oceanology & Marine Fisheries, Jiangsu, Nantong 226007, China; 2. Department of Biology and Food Engineering, Changshu Institute of Technology, Changshu 215000, China)
Oct., 27, 2009
Exopalaemon carinicauda; feeding behavior; cannibalism; defence behavior
This study described the behaviors of feeding, cannibalism, locomotion, molting, spacing, attacking and defence of Exopalaemon carinicauda cultured in high-density farming system. The results revealed that feeding behavior was active; locomotion behavior was diversiform; cannibalism behavior was obviously related to both of starvation and variability of the shrimp body size and the rate of cannibalism would increase with the increase of time separation of feeding and the increase of rearing density; the molting behavior was special, which occurred several times during a molting process; and spacing behavior was not obviously and the frequency of attacking was low.
S968.22
A
1000-3096(2010)10-0053-04
2009-10-27;
2010-03-16
国家海洋局海洋公益性行业科研专项课题资助项目(200705009); 江苏省社会发展科技计划资助项目(BS2006029)
沈辉(1981-), 男, 江苏徐州人, 助理研究员, 主要从事海洋生物学研究, E-mail: darkhui@163.com
(本文编辑: 谭雪静)
